- Theories of general anaesthetic action
-
 Structures of general anaesthetics widely used in medicine.
Structures of general anaesthetics widely used in medicine.
A general anaesthetic (or anesthetic) is a drug that brings about a reversible loss of consciousness. These drugs are generally administered by an anaesthetist/anaesthesiologist in order to induce or maintain general anaesthesia to facilitate surgery. General anaesthetics have been widely used in surgery since 1842 when Crawford Long for the first time administered diethyl ether to a patient and performed a painless operation. It has always been believed that general anaesthetics exert their effects (analgesia, amnesia, immobility) by modulating the activity of membrane proteins in the neuronal membrane. However, the exact location and mechanism of this action are still largely unknown although much research has been done in this area. There are a number of outdated and modern theories that attempt to explain anaesthetic action.
The concept of specific interactions between receptors and drugs first introduced by Paul Ehrlich [1] states that drugs act only when they are bound to their targets (receptors). However, this concept is not working well in case of general anaesthetics because:
- Molecular structures of general anaesthetics widely used in medicine are very simple and diverse so that there is no obvious structure–activity relationship [1] (see structures of general anaesthetics widely used in medicine: 1-ethanol, 2- chloroform, 3-diethylether, 4-fluroxene, 5-halothane, 6-metheoxyflurane, 7- enflurane, 8-isoflurane, 9-desflurane, 10-sevoflurane)
- Most general anaesthetics have remarkably weak affinity for their targets acting at much higher concentrations than most other drugs so that diverse side effects are inevitable [ref].
All these common features of general anaesthetics made it hard for early researchers to believe that general anaesthetics act in a specific manner and their action on neuronal membrane was thought to be global (through nonspecific perturbation of lipid membrane of CNS neurons) rather than through specific sites.
Contents
Lipid solubility-anaesthetic potency correlation (the Meyer-Overton correlation)
 The Meyer-Overton correlation for anaesthetics
The Meyer-Overton correlation for anaestheticsThe nonspecific mechanism of general anaesthetic action was first proposed by Von Bibra and Harless in 1847 .[2] They suggested that general anaesthetics may act by dissolving in the fatty fraction of brain cells and removing fatty constituents from them, thus changing activity of brain cells and inducing anaesthesia. In 1899 Hans Horst Meyer published the first experimental evidence of the fact that anaesthetic potency is related to lipid solubility in his article entitled "Zur Theorie der Alkoholnarkose" [3] [4] .[5] Two years later a similar theory was published independently by Overton .[6]
Meyer compared the potency of many agents, defined as the reciprocal of the molar concentration required to induce anaesthesia in tadpoles, with their olive oil/water partition coefficient. He found a nearly linear relationship between potency and the partition coefficient for many types of anaesthetic molecules such as alcohols, aldehydes, ketones, ethers, and esters. The anaesthetic concentration required to induce anaesthesia in 50% of a population of animals (the EC50) was independent of the means by which the anaesthetic was delivered, i.e., the gas or aqueous phase [3] [4] [5] .[7]
Meyer and Overton had discovered the striking correlation between the physical properties of general anaesthetic molecules and their potency: the greater is the lipid solubility of the compound in olive oil the greater is its anaesthetic potency .[7] This correlation is true for a wide range of anaesthetics with lipid solubilities ranging over 4-5 orders of magnitude if olive oil is used as the oil phase. However, this correlation can be improved considerably in terms of both the quality of the correlation and the increased range of anaesthetics if bulk octanol [8] or a fully hydrated fluid lipid bilayer [9] [10] [11] [12] is used as the “oil” phase. It was noted also that volatile anaesthetics are additive in their effects (a mixture of a half dose of two different volatile anaesthetics gave the same anaesthetic effect as a full dose of either drug alone).
Nb: The y-axis in the accompanying graph is incorrectly labelled and should read "log of Minimum Alveolar Concentration or MAC" - The lesser the MAC the greater the potency (Halothane has a MAC of slightly less than 1 while Nitrous oxide has a MAC of around 105, halothane is much more potent and it is because the log of the MAC is plotted on the y axis that halothane has a value of 0.01 and nitrous oxide has a value of 1. The drug potency increases and the dose required to produce anaesthesia reduces as the oil:gas solubility increases.
Outdated lipid hypotheses of general anaesthetic action
 Bulky and hydrophobic anaesthetic molecules accumulate inside the neuronal cell membrane causing its distortion and expansion (thickening) due to volume displacement. Membrane thickening reversibly alters function of membrane ion channels thus providing anaesthetic effect. Actual chemical structure of the anaesthetic agent per se was not important, but its molecular volume plays the major role: the more space within membrane is occupied by anaesthetic - the greater is the anaesthetic effect.
Bulky and hydrophobic anaesthetic molecules accumulate inside the neuronal cell membrane causing its distortion and expansion (thickening) due to volume displacement. Membrane thickening reversibly alters function of membrane ion channels thus providing anaesthetic effect. Actual chemical structure of the anaesthetic agent per se was not important, but its molecular volume plays the major role: the more space within membrane is occupied by anaesthetic - the greater is the anaesthetic effect.From the correlation between lipid solubility and anaesthetic potency, both Meyer and Overton had surmised a unitary mechanism of general anaesthesia. They assumed that solubilization of lipophilic general anaesthetic in lipid bilayer of the neuron causes its malfunction and anaesthetic effect when critical concentration of anaesthetic is reached. Later in 1973 Miller and Smith suggested the critical volume hypothesis also called lipid bilayer expansion hypothesis.[13] They assumed that bulky and hydrophobic anaesthetic molecules accumulate inside the hydrophobic (or lipophilic) regions of neuronal lipid membrane causing its distortion and expansion (thickening) due to volume displacement. Accumulation of critical amounts of anaesthetic causes membrane thickening sufficient to reversibly alter function of membrane ion channels thus providing anaesthetic effect. Actual chemical structure of the anaesthetic agent per se is not important, but its molecular volume plays the major role: the more space within membrane is occupied by anaesthetic - the greater is the anaesthetic effect. Based on this theory, in 1954 Mullins suggested that the Meyer-Overton correlation with potency can be improved if molecular volumes of anaesthetic molecules are taken into account .[14] This theory existed for over 60 years and was supported by experimental fact that increases in atmospheric pressure reverse anaesthetic effect (pressure reversal effect).
Then other theories of anaesthetic action emerged mostly ‘physicochemical’ theories that took into account the diverse chemical nature of general anaesthetics and suggested that anaesthetic effect is exerted through some perturbation of the lipid bilayer. Several types of bilayer perturbations were proposed to cause anaesthetic effect (reviews [15] [16] .[17] ):
- changes in phase separation
- changes in bilayer thickness
- changes in order parameters
- changes in curvature elasticity
According to the lateral phase separation theory [18] anaesthetics exert their action by fluidizing nerve membranes to a point when phase separations in the critical lipid regions disappear. This anaesthetic-induced fluidization makes membranes less able to facilitate the conformational changes in proteins that may be the basis for such membrane events as ion gating, synaptic transmitter release, and transmitter binding to receptors.
All these outdated lipid theories generally suffer from four weaknesses [1] (full description see in sections below):
- Stereoisomers of an anaesthetic drug have very different anaesthetic potency whereas their oil/gas partition coefficients are similar
- Certain highly lipid soluble drugs expected to act as anaesthetics, exert convulsive effect instead of anaesthetic effect and therefore such drugs were termed nonimmobilizers.
- General anaesthetic-induced changes in membrane density and fluidity are so small that relatively small increases in temperature can mimic these effects on membrane fluidity and density without causing anaesthesia.
- Cutoff effect - addition of methylene groups to a homologous series of long chain alcohols, or alkanes, increases their lipid solubility and should thereby produce a corresponding increase in anaesthetic potency, but it was observed only to a certain "cutoff" length of the chain.
In conclusion it is important to point out that the correlation between lipid solubility and potency of general anaesthetics is a necessary but not sufficient condition for inferring a lipid target site. General anaesthetics could equally well be binding to hydrophobic target sites on proteins in the brain. The main reason that more polar general anaesthetics are less potent is that they have to cross the blood-brain barrier to exert their effect on neurons in the brain.
Objections to the outdated lipid hypotheses
1. Stereoisomers of an anaesthetic drug
Stereoisomers that represent mirror images of each other are termed enantiomers or optical isomers (for example, the isomers of R-(+)- and S-(-)-etomidate) .[1] Physicochemical effects of enantiomers are always identical in an achiral environment (for example in the lipid bilayer). However, in vivo enantiomers of many general anaesthetics (e.g. isoflurane, thiopental, etomidate) can differ greatly in their anaesthetic potency despite the similar oil/gas partition coefficients [19] [20] For example, the R-(+) isomer of etomidate is 10 times more potent anaesthetic than its S-(-) isomer .[1] This means that optical isomers partition identically into lipid, but have differential effects on ion channels and synaptic transmission. This objection provides a compelling evidence that the primary target for anaesthetics is not the achiral lipid bilayer itself but rather stereoselective binding sites on membrane proteins that provide a chiral environment for specific anaesthetic-protein docking interactions .[1]
2. Nonimmobilizers
All general anaesthetics induce immobilization (absence of movement in response to noxious stimuli) through depression of spinal cord functions, whereas their amnesic actions are exerted within the brain. According to the Meyer-Overton correlation the anaesthetic potency of the drug is directly proportional to its lipid solubility, however, there are many compounds that do not satisfy this rule. These drugs are strikingly similar to potent general anaesthetics and are predicted to be potent anaesthetics based on their lipid solubility, but they exert only one constituent of the anaesthetic action (amnesia) and do not suppress movement (i.e. do not depress spinal cord functions) as all anaesthetics do [21] [22] [23] .[24] These drugs are referred to as nonimmobilizers. The existence of nonimmobilizers suggests that anaesthetics induce different components of anaesthetic effect (amnesia and immobility) by affecting different molecular targets and not just the one target (neuronal bilayer) as it was believed earlier .[25] Good example of immobilizers are halogenated alkanes that are very hydrophobic, but fail to suppress movement in response to noxious stimulation at appropriate concentrations.
3. General anaesthetic-induced changes in membrane density and fluidity
Experimental studies have shown that general anaesthetics including ethanol are potent fluidizers of natural and artificial membranes. However, changes in membrane density and fluidity in the presence of clinical concentrations of general anaesthetics are so small that relatively small increases in temperature (~1°C) can mimic them without causing anaesthesia .[26] The change in body temperature of approximately 1°C is within the physiological range and clearly it is not sufficient to induce loss of consciousness per se. Thus membranes are fluidized only by large quantities of anaesthetics, but there are no changes in membrane fluidity when concentrations of anaesthetics are small and restricted to pharmacologically relevant.
4. Cutoff effect
According to the Meyer-Overton correlation, the addition of methylene groups to a homologous series of any general anaesthetic (e.g. n-alcohols, or alkanes) increases their lipid solubility and thereby should produce a corresponding increase in anaesthetic potency. However, addition of methylene groups cannot make anaesthetic increasingly potent without limit and at a certain chain length (cutoff chain length) the addition of just one methylene group causes the molecule to lose its ability to anaesthetise. For the n-alcohols the cutoff occurs at a carbon chain length of about 13 [27] and for the n-alkanes at a chain length of between 6 and 10, depending on the species .[28]
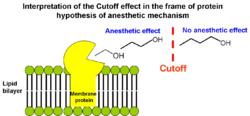 The Meyer-Overton rule predicts the constant increase of anaesthetic potency in the raw of n-alkanols with the increase of n-alkanol chain length. However, n-alkanols with chain length above certain “cutoff” length surprisingly lack anaesthetic effect because their hydrocarbon chain volume starts to exceed volume of the putative binding site on the protein.
The Meyer-Overton rule predicts the constant increase of anaesthetic potency in the raw of n-alkanols with the increase of n-alkanol chain length. However, n-alkanols with chain length above certain “cutoff” length surprisingly lack anaesthetic effect because their hydrocarbon chain volume starts to exceed volume of the putative binding site on the protein.If general anaesthetics disrupt ion channels by partitioning into and perturbing the lipid bilayer, then one would expect that their solubility in lipid bilayers would also display the cutoff effect. However, partitioning of alcohols into lipid bilayers does not display a cutoff for long-chain alcohols from n-decanol to n-pentadecanol. A plot of chain length vs. the logarithm of the lipid bilayer/buffer partition coefficient K is linear, with the addition of each methylene group causing a change in the Gibbs free energy of -3.63 kJ/mol.
The cutoff effect was first interpreted as evidence that anaesthetics exert their effect not by acting globally on membrane lipids but rather by binding directly to hydrophobic pockets of well-defined volumes in proteins. As the alkyl chain grows, the anaesthetic fills more of the hydrophobic pocket and binds with greater affinity. When the molecule is too large to be entirely accommodated by the hydrophobic pocket, the binding affinity no longer increases with increasing chain length. Thus the volume of the n-alkanol chain at the cutoff length provides an estimate of the binding site volume. This objection provided the basis for protein hypothesis of anaesthetic effect (see below).
 A) Short hydrocarbon chains are relatively rigid in terms of conformational enthropy and are close to alkanol hydroxyl group (“buoy”) tethered to the interface. This makes short chain alkanols efficient mediators that redistribute lateral stress from membrane interior to its interface. B) This ability decreases in the row of n-alkanols since longer chains are more flexible and are not so tightly tethered to the hydroxyl group. C) Polyhydroxyalkanes 1,6,11,16-hexadecanetetraol and 2,7,12,17-octadecanetetraol exhibit significant anaesthetic potency as was predicted by cutoff effect because the length of the hydrocarbon chain between hydroxyl groups is smaller than the cutoff.
A) Short hydrocarbon chains are relatively rigid in terms of conformational enthropy and are close to alkanol hydroxyl group (“buoy”) tethered to the interface. This makes short chain alkanols efficient mediators that redistribute lateral stress from membrane interior to its interface. B) This ability decreases in the row of n-alkanols since longer chains are more flexible and are not so tightly tethered to the hydroxyl group. C) Polyhydroxyalkanes 1,6,11,16-hexadecanetetraol and 2,7,12,17-octadecanetetraol exhibit significant anaesthetic potency as was predicted by cutoff effect because the length of the hydrocarbon chain between hydroxyl groups is smaller than the cutoff.However, cutoff effect can still be explained in the frame of lipid hypothesis [29] .[30] In short-chain alkanols (A) segments of the chain are rather rigid (in terms of conformational enthropy) and very close to hydroxyl group tethered to aqueous interfacial region ("buoy"). Consequently, these segments efficiently redistribute lateral stresses from the bilayer interior toward the interface. In long-chain alkanols (B) hydrocarbon chain segments are located further from hydroxyl group and are more flexible than in short-chain alkanols. Efficiency of pressure redistribution decreases as the length of hydrocarbon chain increases until anaesthetic potency is lost at some point. It was proposed that polyalkanols (C) will have anaesthetic effect similar to short-chain 1-alkanols if the chain length between two neighbouring hydroxyl groups is smaller than the cutoff .[31] This idea was supported by the experimental evidence because polyhydroxyalkanes 1,6,11,16-hexadecanetetraol and 2,7,12,17-octadecanetetraol exhibited significant anaesthetic potency as was originally proposed .[30]
Modern lipid hypothesis
 General anaesthetic changes membrane lateral pressure profile which determines conformation of membrane ion channel (green lock)
General anaesthetic changes membrane lateral pressure profile which determines conformation of membrane ion channel (green lock)Modern version of lipid hypothesis states that anaesthetic effect happens if solubilization of general anaesthetic in the bilayer causes redistribution of membrane lateral pressures [29] .[32]
Each bilayer membrane has distinct profile of how lateral pressures are distributed within it. Most membrane proteins especially ion channels are sensitive to changes in this lateral pressure distribution profile. These lateral stresses are rather large and vary with depth within the membrane. According to modern lipid hypothesis change in membrane lateral pressure profile shifts the conformational equilibrium of certain membrane proteins known to be affected by clinical concentrations of anaesthetics such as ligand-gated ion channels. This mechanism is also nonspecific because potency of the anaesthetic is determined not by its actual chemical structure, but by positional and orientational distribution of its segments and bonds of within the bilayer. However, it was not obvious what is the exact molecular mechanism... Detailed mechanism of general anaesthesia was suggested and investigated using lattice statistical thermodynamics.[32] It was proposed that incorporation of amphiphilic and other interfacially active solutes (e.g. general anaesthetics) into the bilayer increases the lateral pressure selectively near the aqueous interfaces, which is compensated by decrease in lateral pressure toward the centre of the bilayer. Calculations showed that general anaesthesia likely involves inhibition of the opening of the ion channel in a postsynaptic ligand-gated membrane protein [32] by the following mechanism:
- Channel is trying to open in the response to nerve impulse thus increasing the cross-sectional area of the protein more near the aqueous interface than in the middle of the bilayer,
- Then the anaesthetic-induced increase in lateral pressure near the interface shifts the protein conformational equilibrium to back to the closed state, since channel opening will require greater work against the higher pressure at interface. This is the first hypothesis that provided not just correlations of potency with structural or thermodynamic properties, but a detailed mechanistic and thermodynamic understanding of anaesthesia.
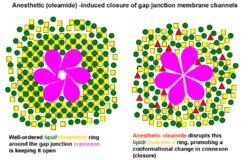
Thus according to the modern lipid hypothesis anaesthetics do not act directly on their membrane protein targets, but rather perturb specialized lipid matrices at the protein-lipid interface, which act as mediators. This is a new kind of transduction mechanism, different from the usual key-lock interaction of ligand and receptor, where anaesthetic (ligand) affects the function of membrane proteins by binding to the specific site on the protein. Thus some membrane proteins are proposed to be sensitive to their lipid environment. Slightly different detailed molecular mechanism of how bilayer perturbation can influence the ion-channel was proposed in the same year. Oleamide (fatty acid amide of oleic acid) is an endogenous anaesthetic found in vivo (in the cat’s brain) and it is known to potentiate sleep and lower temperature of the body through closing gap junction channel connexon .[33] The detailed mechanism is shown on the picture: the well ordered lipid(green)/cholesterol(yellow) ring that exists around connexon (magenta) becomes disordered on treatment with anaesthetic (red triangles), promoting a closure of connexon ion channel. This decreases brain activity and induces lethargy and anaesthetic effect.
Membrane protein hypothesis of general anaesthetic action
In the early 1980s, Franks and Lieb [34] demonstrated that the Meyer-Overton correlation can be reproduced using a soluble protein. They found that two classes of proteins are inactivated by clinical doses of anaesthetic in the total absence of lipid. These are luciferases, which are used by bioluminescent animals and bacteria to produce light, [35] and cytochrome P450 [36] , which is a group of heme proteins that hydroxylate a diverse group of compounds, including fatty acids, steroids, and xenobiotics such as phenobarbital. Remarkably, inhibition of these proteins by general anaesthetics was directly correlated with their anaesthetic potencies. Luciferase inhibition also exhibits a long-chain alcohol cutoff, which is related to the size of the anaesthetic-binding pocket .[37]
These observations were important because they demonstrated that general anaesthetics may also interact with hydrophobic protein sites of certain proteins, rather than affect membrane proteins indirectly through nonspecific interactions with lipid bilayer as mediator [8] .[38] It was shown that anaesthetics alter the functions of many cytoplasmic signalling proteins, including protein kinase C, [39] [40] however, the proteins considered the most likely molecular targets of anaesthetics are ion channels. According to this theory general anaesthetics are much more selective than in the frame of lipid hypothesis and they bind directly only to small number of targets in CNS mostly ligand(neurotransmitter)-gated ion channels in synapse and G-protein coupled receptors altering their ion flux. Particularly Cys-loop receptors [41] are plausible targets for general anaesthetics that bind at the interface between the subunits. The Cys-loop receptor superfamily includes inhibitory receptors (GABA A, GABA C, glycine receptors) and excitatory receptors (acetylcholine receptor and 5-HT3 serotonin receptor). General anaesthetics can inhibit the channel functions of excitatory receptors or potentiate functions of inhibitory receptors, respectively. Although protein targets for anaesthetics have been partly identified the exact nature of general anaesthetic-protein interactions still remains a mystery.
It was initially hypothesized that general anaesthetic binds to its target ion channel by a key-lock mechanism and changes its structure dramatically from open to closed conformation or vice versa. However, there is lots of evidence accumulated against direct key-lock interaction of membrane proteins with general anaesthetics [42] [43] [44] .[45]
 Inhaled general anaesthetics frequently do not change structure of their target protein (of Cys-loop receptor here) but change its dynamics especially dynamics in the flexible loops that connect α-helices in a bundle thus disrupting modes of motion essential for the protein function.
Inhaled general anaesthetics frequently do not change structure of their target protein (of Cys-loop receptor here) but change its dynamics especially dynamics in the flexible loops that connect α-helices in a bundle thus disrupting modes of motion essential for the protein function.Various studies have shown that low affinity drugs including inhaled general anaesthetics do not usually interact with their target proteins via specific lock-and-key binding mechanism because they do not change molecular structures of transmembrane receptors, ion channels and globular proteins. Based on these experimental facts and some computer simulations modern version of protein hypothesis was proposed [46] .[47] Proteins of four-α-helix bundle structural motif served as models of monomer of pentameric Cys-loop receptor because binding pockets of inhaled anaesthetics are believed to be within transmembrane four-α-helix bundles of Cys-loop receptors .[48] Inhaled general anaesthetic does not change structure of membrane channel but changes its dynamics especially dynamics in the flexible loops that connect α-helices in a bundle and are exposed to the membrane-water interface. It is a well known fact that dynamics of protein in microsecond-millisecond timescale is often coupled with functions of the protein. Thus it was logical to propose that since inhaled general anaesthetics do not change protein structure they may exert their effect on proteins by modulating protein dynamics in a slow microsecond-millisecond timescale and/or by disrupting the modes of motion essential for function of this protein. Normal interactions between residues in protein regions (loops) at the water-lipid interface that play critical roles in protein functions and agonist binding may be disrupted by general anaesthetic. Interactions within the same loop or between different loops may be disrupted by anaesthetics and ultimately functions of Cys-loop receptors may be altered.
References
- ^ a b c d e f Cameron JW (January 2006). "The molecular mechanisms of general anaesthesia: dissecting the GABAA receptor". Continuing Education in Anaesthesia, Critical Care & Pain 6 (2): 49–53. doi:10.1093/bjaceaccp/mki068. http://ceaccp.oxfordjournals.org/cgi/content/abstract/mki068v1.
- ^ Harless E, von Bibra E (1847). "Die Ergebnisse der Versuche über die Wirkung des Schwefeläthers". Erlangen.
- ^ a b Meyer HH (1899). "Zur Theorie der Alkoholnarkose". Arch. Exp. Pathol. Pharmacol. 42 (2–4): 109–118. doi:10.1007/BF01834479.
- ^ a b Meyer HH (1899). "Welche eigenschaft der anasthetica bedingt inre Narkotische wirkung?". Arch. Exp. Pathol. Pharmakol. 42 (2–4): 109–118. doi:10.1007/BF01834479.
- ^ a b Meyer HH (1901). "Zur Theorie der Alkoholnarkose. Der Einfluss wechselnder Temperature auf Wirkungsstärke und Theilungscoefficient der Narcotica". Arch. Exp. Pathol. Pharmakol. 46 (5–6): 338–346. doi:10.1007/BF01978064.
- ^ Overton CE (1901). "Studien über die Narkose zugleich ein Beitrag zur allgemeinen Pharmakologie". Gustav Fischer, Jena, Switzerland.
- ^ a b Meyer KH (1937). "Contributions to the theory of narcosis". Trans Faraday Soc 33: 1062–8. doi:10.1039/tf9373301062.
- ^ a b Franks NP, Lieb WR (July 1978). "Where do general anesthetics act?". Nature 274 (5669): 339–42. doi:10.1038/274339a0. PMID 672957. http://www.nature.com/nature/journal/v274/n5669/pdf/274339a0.pdf.
- ^ Janoff AS, Pringle MJ, Miller KW (1981). "Correlation of general anesthetic potency with solubility in membranes". Biochim. Biophys. Acta 649 (1): 125–8. doi:10.1016/0005-2736(81)90017-1. PMID 7306543. http://www.sciencedirect.com/science?_ob=MiamiImageURL&_imagekey=B6T1T-47TG6GB-CC-1&_cdi=4899&_user=2139813&_check=y&_orig=search&_coverDate=11%2F20%2F1981&view=c&wchp=dGLzVtb-zSkzk&md5=a91d93fe0805e51d8a3c9fa53fd64700&ie=/sdarticle.pdf.
- ^ Taheri S, Halsey MJ, Liu J, Eger EI, Koblin DD, Laster MJ (1991). "What solvent best represents the site of action of inhaled anesthetics in humans, rats, and dogs?". Anesth. Analg. 72 (5): 627–34. doi:10.1213/00000539-199105000-00010. PMID 2018220. http://www.anesthesia-analgesia.org/cgi/content/abstract/72/5/627.[dead link]
- ^ Vaes WHJ, Ramos EU, Hamwijk C, van Holsteijn I, Blaauboer BJ, Seinen W, Verhaar HJM, Hermens JLM (1997). "Solid phase microextraction as a tool to determine membrane/water partition coefficients and bioavailable concentrations in in vitro systems". Chem. Res. Toxicol. 10 (10): 1067–72. doi:10.1021/tx970109t. PMID 9348427. http://pubs.acs.org/doi/abs/10.1021/tx970109t.
- ^ Meijer LA, Leermakers FAM, Lyklema J (1999). "Self-consistent-field modeling of complex molecules with united atom detail in inhomogeneous systems. cyclic and branched foreign molecules in dimyristoylphosphatidylcholine membranes". J. Chem. Phys. 110 (6560): 6560–79. doi:10.1063/1.478562. http://scitation.aip.org/getabs/servlet/GetabsServlet?prog=normal&id=JCPSA6000110000013006560000001&idtype=cvips&gifs=yes.
- ^ Miller KW, Paton WD, Smith RA, Smith EB (March 1973). "The pressure reversal of general anesthesia and the critical volume hypothesis". Mol Pharmacol. 9 (2): 131–43. PMID 4711696.
- ^ Mullins LI (1954). "Some physical mechanisms in narcosis". Chem. Rev. 54 (2): 289–323. doi:10.1021/cr60168a003.
- ^ Miller KW (1985). "The nature of the site of general anesthesia". International review of neurobiology. International Review of Neurobiology 27 (1): 1–61. doi:10.1016/S0074-7742(08)60555-3. ISBN 9780123668271. PMID 3910602.
- ^ Janoff AS, Miller KW (1982). "A critical assessment of the lipid theories of general anaesthetic action". Biological Membranes 4 (1): 417–76.
- ^ Koblin DD (1994). "Inhaled anesthetics: mechanisms of action". Anesthesia 4th edition: 67–99.
- ^ Trudell JR (1977). "A unitary theory of anesthesia based on lateral phase separations in nerve membranes". Anesthesiology 46 (1): 5–10. doi:10.1097/00000542-197701000-00003. PMID 12686.
- ^ Nau C, Strichartz GR (August 2002). "Drug chirality in anesthesia". Anesthesiology 97 (2): 497–502. doi:10.1097/00000542-200208000-00029. PMID 12151942.
- ^ Franks NP, Lieb WR (October 1991). "Stereospecific effects of inhalational general anesthetic optical isomers on nerve ion channels". Science 254 (5030): 427–30. doi:10.1126/science.1925602. PMID 1925602. http://www.sciencemag.org/cgi/content/abstract/254/5030/427.
- ^ Kandel L, Chortkoff BS, Sonner J, Laster MJ, Eger EI (1996). "Nonanesthetics can suppress learning". Anesth Analg 82 (2): 321–6. doi:10.1097/00000539-199602000-00019. PMID 8561335. http://www.anesthesia-analgesia.org/cgi/content/abstract/82/2/321.
- ^ Koblin DD, Chortkoff BS, Laster MJ, Eger EI II, Halsey MJ, Ionescu P (1994). "Polyhalogenated and perfluorinated compounds that disobey the Meyer-Overton hypothesis". Anesth Analg 79 (6): 1043–8. doi:10.1213/00000539-199412000-00004. PMID 7978424. http://www.anesthesia-analgesia.org/cgi/content/abstract/79/6/1043.
- ^ Fang Z, Sonner J, Laster MJ, Ionescu P, Kandel L, Koblin DD, Eger EI II, Halsey MJ (1996). "Anesthetic and convulsant properties of aromatic compounds and cycloalkanes: Implications for mechanisms of narcosis". Anesth. Analg. 83 (5): 1097–1104. doi:10.1097/00000539-199611000-00035. PMID 8895293. http://www.anesthesia-analgesia.org/cgi/content/abstract/83/5/1097.
- ^ Taheri S, Laster MJ, Liu J, Eger EI II, Halsey MJ, Koblin DD (1993). "Anesthesia by n-alkanes not consistent with the Meyer-Overton hypothesis: Determinations of solubilities of alkanes in saline and various lipids". Anesth Analg 77 (1): 7–11. doi:10.1213/00000539-199307000-00003. PMID 8317750. http://www.anesthesia-analgesia.org/cgi/content/abstract/77/1/7?ijkey=6421e455804cc9b44943b9c1ed998f620fb65e6b&keytype2=tf_ipsecsha.
- ^ Eger E, Koblin D, Harris R, et al. (1997). "Hypothesis: Inhaled anesthetics produce immobility and amnesia by different mechanisms at different sites". Anesth Analg. 84 (4): 915–918. doi:10.1097/00000539-199704000-00039. PMID 9085981. http://www.anesthesia-analgesia.org/cgi/reprint/84/4/915.pdf.
- ^ Franks NP, Lieb WR (December 1982). "Molecular mechanisms of general anaesthesia". Nature 300 (9): 487–93. doi:10.1038/300487a0. http://www.nature.com/nature/journal/v300/n5892/pdf/300487a0.pdf.
- ^ Pringle MJ, Brown KB, Miller KW (1981). "Can the Lipid Theories of Anesthesia Account for the Cutoff in Anesthetic Potency in Homologous Series of Alcohols?". Mol. Pharmacol. 19 (1): 49–55. PMID 7207463. http://molpharm.aspetjournals.org/cgi/reprint/19/1/49.
- ^ Liu J, Laster MJ, Taheri S, Eger EI, Koblin DD, Halsey MJ (1993). "Is There a Cutoff in Anesthetic Potency for the Normal Alkanes?". MAnesth Analg 77 (1): 12–18. PMID 8317717. http://www.anesthesia-analgesia.org/cgi/content/abstract/77/1/12.[dead link]
- ^ a b Eckenhoff RG, Tanner JW, Johansson JS (1 August 1999). "Steric hindrance is not required for n-alkanol cutoff in soluble proteins". Mol. Pharmacol. 56 (2): 414–8. PMID 10419562. http://molpharm.aspetjournals.org/cgi/content/full/56/2/414.
- ^ a b Mohr JT, Gribble GW, Lin SS, Eckenhoff RG, Cantor RS (April 2005). "Anesthetic Potency of Two Novel Synthetic Polyhydric Alkanols Longer than the n-Alkanol Cutoff: Evidence for a Bilayer-Mediated Mechanism of Anesthesia?". J. Med. Chem. 48 (12): 4172–76. doi:10.1021/jm049459k. PMID 15943489. http://pubs.acs.org/doi/abs/10.1021/jm049459k.
- ^ Cantor RS (May 2001). "Breaking the Meyer-Overton rule: predicted effects of varying stiffness and interfacial activity on the intrinsic potency of anesthetics". Biophys. J. 80 (5): 2284–97. Bibcode 2001BpJ....80.2284C. doi:10.1016/S0006-3495(01)76200-5. PMC 1301419. PMID 11325730. http://linkinghub.elsevier.com/retrieve/pii/S0006-3495(01)76200-5.
- ^ a b c Cantor RS (December 1997). "The Lateral Pressure Profile in Membranes: A Physical Mechanism of General Anesthesia". Biochemistry 36 (9): 2339–44. doi:10.1021/bi9627323. PMID 9054538. http://pubs.acs.org/doi/abs/10.1021/bi9627323.
- ^ Lerner RA (December 1997). "A hypothesis about the endogenous analogue of general anesthesia". Proc. Natl. Acad. Sci. USA 94 (25): 13375–7. doi:10.1073/pnas.94.25.13375. PMC 33784. PMID 9391028. http://www.pnas.org/content/94/25/13375.full?ck=nck.
- ^ Franks NP, Lieb WR (August 1984). "Do general anaesthetics act by competitive binding to specific receptors?". Nature 310 (16): 599–601. doi:10.1038/310599a0. PMID 6462249. http://www.nature.com/nature/journal/v310/n5978/pdf/310599a0.pdf.
- ^ Franks NP, Jenkins A, Conti E, Lieb WR, Brick P (November 1998). "Structural basis for the inhibition of firefly luciferase by a general anesthetic". Biophys. J. 75 (5): 2205–11. Bibcode 1998BpJ....75.2205F. doi:10.1016/S0006-3495(98)77664-7. PMC 1299894. PMID 9788915. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1299894.
- ^ LaBella FS, Stein D, Queen G (October 1998). "Occupation of the cytochrome P450 substrate pocket by diverse compounds at general anesthesia concentrations". European Journal of Pharmacology 358 (2): 177–85. doi:10.1016/S0014-2999(98)00596-2. PMID 9808268. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6T1J-3TVP8HH-C&_user=2139813&_rdoc=1&_fmt=&_orig=search&_sort=d&view=c&_acct=C000054276&_version=1&_urlVersion=0&_userid=2139813&md5=76af9e3fb90bac44d8281e0b1138971a.
- ^ Franks NP, Lieb WR (July 1985). "Mapping of general anesthetic target sites provides a molecular basis for cutoff effects". Nature 316 (6026): 349–51. doi:10.1038/316349a0. PMID 4022125. http://www.nature.com/nature/journal/v316/n6026/abs/316349a0.html.
- ^ Miller KW (1985). "The nature of the site of general anesthesia". Int Rev Neurobiol. International Review of Neurobiology 27: 1–61. doi:10.1016/S0074-7742(08)60555-3. ISBN 9780123668271. PMID 3910602.
- ^ Slater SJ, Cox KJ, Lombardi JV, et al (July 1993). "Inhibition of protein kinase C by alcohols and anaesthetics". Nature 364 (6432): 82–4. doi:10.1038/364082a0. PMID 8316305. http://www.nature.com/nature/journal/v364/n6432/pdf/364082a0.pdf.
- ^ Hemmings HC Jr, Adamo AI (1994). "Effects of halothane and propofol on purified brain protein kinase C activation". Anesthesiology 81 (1): 147–55. doi:10.1097/00000542-199409001-00886. PMID 8042784. http://journals.lww.com/anesthesiology/pages/articleviewer.aspx?year=1994&issue=07000&article=00021&type=abstract.
- ^ Franks NP, Lieb WR (February 1994). "Molecular and Cellular Mechanisms of General Anesthesia". Nature 367 (17): 607–14. doi:10.1038/367607a0. PMID 7509043. http://www.nature.com/nature/journal/v367/n6464/pdf/367607a0.pdf.
- ^ Johansson JS, Gibney BR, Rabanal F, Reddy KS, Dutton PL (January 1998). "A Designed Cavity in the Hydrophobic Core of a Four-a-Helix Bundle Improves Volatile Anesthetic Binding Affinity". Biochemistry 37 (5): 1421–29. doi:10.1021/bi9721290. PMID 9477971. http://pubs.acs.org/doi/abs/10.1021/bi9721290.
- ^ Cui T, Bondarenko V, Ma D, Canlas C, Brandon NR, Johansson JS, Xu Y, Tang P (2008). "Four-α-Helix Bundle with Designed Anesthetic Binding Pockets. Part II: Halothane Effects on Structure and Dynamics". Biophys. J. 94 (11): 4464–72. doi:10.1529/biophysj.107.117853. PMC 2480694. PMID 18310239. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B94RW-4VB4SVD-14&_user=2139813&_coverDate=06%2F01%2F2008&_fmt=high&_orig=browse&_sort=d&view=c&_acct=C000054276&_version=1&_urlVersion=0&_userid=2139813&md5=470a0c6f2626517c01372bcf65be342a.
- ^ Ma D, Brandon NR, Cui T, Bondarenko V, Canlas C, Johansson JS, Tang P, Xu Y (2008). "Four-α-Helix Bundle with Designed Anesthetic Binding Pockets. Part I: Structural and Dynamical Analyses". Biophys. J. 94 (11): 4454–63. doi:10.1529/biophysj.107.117838. PMC 2480675. PMID 18310240. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B94RW-4VB4SVD-13&_user=2139813&_coverDate=06%2F01%2F2008&_fmt=high&_orig=browse&_sort=d&view=c&_acct=C000054276&_version=1&_urlVersion=0&_userid=2139813&md5=3fc05676089f0520addaf99b63695e21.
- ^ Liu R, Loll PJ, Eckenhoff RG (2005). "Structural basis for high-affinity volatile anesthetic binding in a natural 4-helix bundle protein". FASEB J. 19 (6): 567–76. doi:10.1096/fj.04-3171com. PMID 15791007. http://www.fasebj.org/cgi/content/abstract/fasebj;19/6/567.
- ^ Tang P, Xu Y (December 2002). "Large-scale molecular dynamics simulations of general anesthetic effects on the ion channel in the fully hydrated membrane: The implication of molecular mechanisms of general anesthesia". Proc. Natl. Acad. Sci. U.S.A. 99 (25): 16035–40. doi:10.1073/pnas.252522299. PMC 138560. PMID 12438684. http://www.anes.upmc.edu/research/tanglab/abstracts/Proc%20Natl%20Acad%20Sci%20USA,%2099,%2016035-16040,%20(2002).pdf.
- ^ Canlas CG, Cui T, Li L, Xu Y, Tang P (September 2008). "Anesthetic modulation of protein dynamics: insights from a NMR study". J. Phys. Chem. B 112 (45): 14312–8. doi:10.1021/jp805952w. PMC 2669902. PMID 18821786. http://pubs.acs.org/doi/abs/10.1021/jp805952w.
- ^ Mihic SJ, Ye Q, Wick MJ, Koltchine VV, Krasowski MD, Finn SE, Mascia MP, Valenzuela CF, Hanson KK, Greenblatt EP, Harris RA, Harrison NL (September 1997). "Sites of alcohol and volatile anaesthetic action on GABA(A) and glycine receptors". Nature 389 (6649): 385–9. doi:10.1038/38738. PMID 9311780. http://www.nature.com/nature/journal/v389/n6649/full/389385a0.html.
- v. Hintzenstern U , Petermann H, Schwarz W (November 2001). "Frühe Erlanger Beiträge zur Theorie und Praxis der äther- und Chloroformnarkose : Die tierexperimentellen Untersuchungen von Ernst von Bibra und Emil Harless" (in German). Der Anaesthesist 50 (11): 869–80. doi:10.1007/s001010100220. PMID 11760483. http://www.springerlink.com/content/6k52g0tfnw8ea35l/.
Categories:








Wikimedia Foundation. 2010.