- Tyrannosaurus
-
"T. rex" redirects here. For other uses, see T. rex (disambiguation).
Tyrannosaurus rex
Temporal range: Late Cretaceous, 67–65.5 Ma
Skeleton cast, Royal Tyrell Museum Scientific classification 
Phylum: Chordata Class: Reptilia Superorder: Dinosauria Order: Saurischia Suborder: Theropoda Family: †Tyrannosauridae Tribe: †Tyrannosaurini
Osborn, 1906Genus: †Tyrannosaurus
Osborn, 1905Species: †T. rex Binomial name Tyrannosaurus rex
Osborn, 1905Synonyms Genus synonymy- Manospondylus
Cope, 1892 - Dynamosaurus
Osborn, 1905 - Nanotyrannus?
Bakker, Williams & Currie, 1988 - Stygivenator
Olshevsky, 1995 - Dinotyrannus
Olshevsky, 1995
Species synonymy- Manospondylus gigas
Cope, 1892 - Aublysodon amplus?
Marsh, 1892 - Deinodon amplus?
(Marsh, 1892) - Manospondylus amplus?
(Marsh, 1892) - Stygivenator amplus?
(Marsh, 1892) - Tyrannosaurus amplus?
(Marsh, 1892) - Aublysodon cristatus?
Marsh, 1892 - Deinodon cristatus?
(Marsh, 1892) - Stygivenator cristatus?
(Marsh, 1892) - Dynamosaurus imperiosus
Osborn, 1905 - Tyrannosaurus imperiosus
(Osborn, 1905) - Gorgosaurus lancensis?
Gilmore, 1946 - Albertosaurus lancensis?
(Gilmore, 1946) - Aublysodon lancensis?
(Gilmore, 1946) - Deinodon lancensis?
(Gilmore, 1946) - Nanotyrannus lancensis?
(Gilmore, 1946) - Stygivenator molnari
(Paul, 1988) - Dinotyrannus megagracilis
Olshevsky, 1995
Tyrannosaurus (
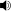 /tɨˌrænɵˈsɔrəs/ or /taɪˌrænɵˈsɔrəs/; meaning "tyrant lizard", from Greek tyrannos (τυράννος) meaning "tyrant," and sauros (σαύρος) meaning "lizard") is a genus of coelurosaurian theropod dinosaur. The species Tyrannosaurus rex (rex meaning "king" in Latin), commonly abbreviated to T. rex, is a fixture in popular culture. It lived throughout what is now western North America, with a much wider range than other tyrannosaurids. Fossils are found in a variety of rock formations dating to the Maastrichtian age of the upper Cretaceous Period, 67 to 65.5 million years ago.[1] It was among the last non-avian dinosaurs to exist before the Cretaceous–Paleogene extinction event.
/tɨˌrænɵˈsɔrəs/ or /taɪˌrænɵˈsɔrəs/; meaning "tyrant lizard", from Greek tyrannos (τυράννος) meaning "tyrant," and sauros (σαύρος) meaning "lizard") is a genus of coelurosaurian theropod dinosaur. The species Tyrannosaurus rex (rex meaning "king" in Latin), commonly abbreviated to T. rex, is a fixture in popular culture. It lived throughout what is now western North America, with a much wider range than other tyrannosaurids. Fossils are found in a variety of rock formations dating to the Maastrichtian age of the upper Cretaceous Period, 67 to 65.5 million years ago.[1] It was among the last non-avian dinosaurs to exist before the Cretaceous–Paleogene extinction event.Like other tyrannosaurids, Tyrannosaurus was a bipedal carnivore with a massive skull balanced by a long, heavy tail. Relative to the large and powerful hindlimbs, Tyrannosaurus forelimbs were small, though unusually powerful for their size, and bore two clawed digits. Although other theropods rivaled or exceeded Tyrannosaurus rex in size, it was the largest known tyrannosaurid and one of the largest known land predators, measuring up to 12.8 m (42 ft) in length,[2] up to 4 metres (13 ft) tall at the hips,[3] and up to 6.8 metric tons (7.5 short tons) in weight.[4] By far the largest carnivore in its environment, Tyrannosaurus rex may have been an apex predator, preying upon hadrosaurs and ceratopsians, although some experts have suggested it was primarily a scavenger. The debate over Tyrannosaurus as apex predator or scavenger is among the longest running in paleontology.
More than 30 specimens of Tyrannosaurus rex have been identified, some of which are nearly complete skeletons. Soft tissue and proteins have been reported in at least one of these specimens. The abundance of fossil material has allowed significant research into many aspects of its biology, including life history and biomechanics. The feeding habits, physiology and potential speed of Tyrannosaurus rex are a few subjects of debate. Its taxonomy is also controversial, with some scientists considering Tarbosaurus bataar from Asia to represent a second species of Tyrannosaurus and others maintaining Tarbosaurus as a separate genus. Several other genera of North American tyrannosaurids have also been synonymized with Tyrannosaurus.
Contents
Description
 Size (in green) compared with selected giant theropods
Size (in green) compared with selected giant theropods
Tyrannosaurus rex was one of the largest land carnivores of all time; the largest complete specimen, FMNH PR2081 ("Sue"), measured 12.8 metres (42 ft) long, and was 4.0 metres (13.1 ft) tall at the hips.[3] Mass estimates have varied widely over the years, from more than 7.2 metric tons (7.9 short tons),[2] to less than 4.5 metric tons (5.0 short tons),[5][6] with most modern estimates ranging between 5.4 and 6.8 metric tons (6.0 and 7.5 short tons).[4][7][8][9] Packard et al. (2009) tested dinosaur mass estimation procedures on elephants and concluded that dinosaur estimations are flawed and produce over-estimations; thus, the weight of Tyrannosaurus could be much less than usually estimated.[10] Other estimations have concluded that the largest known Tyrannosaurus specimens had a weight exceeding 9 tonnes.[11]
Although Tyrannosaurus rex was larger than the well known Jurassic theropod Allosaurus, it was slightly smaller than two other Cretaceous carnivores, Spinosaurus and Giganotosaurus.[12][13]
The neck of Tyrannosaurus rex formed a natural S-shaped curve like that of other theropods, but was short and muscular to support the massive head. The forelimbs had only two clawed fingers,[14] along with an additional small metacarpal representing the remnant of a third digit.[15] In contrast the hind limbs were among the longest in proportion to body size of any theropod. The tail was heavy and long, sometimes containing over forty vertebrae, in order to balance the massive head and torso. To compensate for the immense bulk of the animal, many bones throughout the skeleton were hollow, reducing its weight without significant loss of strength.[14]
 Profile view of a skull (AMNH 5027)
Profile view of a skull (AMNH 5027)The largest known Tyrannosaurus rex skulls measure up to 5 feet (1.5 m) in length.[16] Large fenestrae (openings) in the skull reduced weight and provided areas for muscle attachment, as in all carnivorous theropods. But in other respects Tyrannosaurus's skull was significantly different from those of large non-tyrannosauroid theropods. It was extremely wide at the rear but had a narrow snout, allowing unusually good binocular vision.[17][18] The skull bones were massive and the nasals and some other bones were fused, preventing movement between them; but many were pneumatized (contained a "honeycomb" of tiny air spaces) which may have made the bones more flexible as well as lighter. These and other skull-strengthening features are part of the tyrannosaurid trend towards an increasingly powerful bite, which easily surpassed that of all non-tyrannosaurids.[19][20][21] The tip of the upper jaw was U-shaped (most non-tyrannosauroid carnivores had V-shaped upper jaws), which increased the amount of tissue and bone a tyrannosaur could rip out with one bite, although it also increased the stresses on the front teeth.[22][23]
 Two teeth from the lower jaw of specimen MOR 1125, “B-rex”, showing the variation in tooth size within an individual
Two teeth from the lower jaw of specimen MOR 1125, “B-rex”, showing the variation in tooth size within an individualThe teeth of Tyrannosaurus rex displayed marked heterodonty (differences in shape).[14][24] The premaxillary teeth at the front of the upper jaw were closely packed, D-shaped in cross-section, had reinforcing ridges on the rear surface, were incisiform (their tips were chisel-like blades) and curved backwards. The D-shaped cross-section, reinforcing ridges and backwards curve reduced the risk that the teeth would snap when Tyrannosaurus bit and pulled. The remaining teeth were robust, like "lethal bananas" rather than daggers; more widely spaced and also had reinforcing ridges.[25] Those in the upper jaw were larger than those in all but the rear of the lower jaw. The largest found so far is estimated to have been 30 centimetres (12 in) long including the root when the animal was alive, making it the largest tooth of any carnivorous dinosaur yet found.[3]
Classification
Tyrannosaurus is the type genus of the superfamily Tyrannosauroidea, the family Tyrannosauridae, and the subfamily Tyrannosaurinae; in other words it is the standard by which paleontologists decide whether to include other species in the same group. Other members of the tyrannosaurine subfamily include the North American Daspletosaurus and the Asian Tarbosaurus,[26][27] both of which have occasionally been synonymized with Tyrannosaurus.[23] Tyrannosaurids were once commonly thought to be descendants of earlier large theropods such as megalosaurs and carnosaurs, although more recently they were reclassified with the generally smaller coelurosaurs.[22]
 Reconstructed head and neck in the Naturhistorisches Museum in Vienna
Reconstructed head and neck in the Naturhistorisches Museum in ViennaIn 1955, Soviet paleontologist Evgeny Maleev named a new species, Tyrannosaurus bataar, from Mongolia.[28] By 1965, this species had been renamed Tarbosaurus bataar.[29] Despite the renaming, many phylogenetic analyses have found Tarbosaurus bataar to be the sister taxon of Tyrannosaurus rex,[27] and it has often been considered an Asian species of Tyrannosaurus.[22][30][31] A recent redescription of the skull of Tarbosaurus bataar has shown that it was much narrower than that of Tyrannosaurus rex and that during a bite, the distribution of stress in the skull would have been very different, closer to that of Alioramus, another Asian tyrannosaur.[32] A related cladistic analysis found that Alioramus, not Tyrannosaurus, was the sister taxon of Tarbosaurus, which, if true, would suggest that Tarbosaurus and Tyrannosaurus should remain separate.[26]
Other tyrannosaurid fossils found in the same formations as Tyrannosaurus rex were originally classified as separate taxa, including Aublysodon and Albertosaurus megagracilis,[23] the latter being named Dinotyrannus megagracilis in 1995.[33] However, these fossils are now universally considered to belong to juvenile Tyrannosaurus rex.[34] A small but nearly complete skull from Montana, 60 centimetres (2.0 ft) long, may be an exception. This skull was originally classified as a species of Gorgosaurus (G. lancensis) by Charles W. Gilmore in 1946,[35] but was later referred to a new genus, Nanotyrannus.[36] Opinions remain divided on the validity of N. lancensis. Many paleontologists consider the skull to belong to a juvenile Tyrannosaurus rex.[37] There are minor differences between the two species, including the higher number of teeth in N. lancensis, which lead some scientists to recommend keeping the two genera separate until further research or discoveries clarify the situation.[27][38]
Manospondylus
 Skull of the type specimen (CM 9380) at the Carnegie Museum of Natural History. This was heavily and inaccurately restored with plaster using Allosaurus as a model before it was reverted to its original state.
Skull of the type specimen (CM 9380) at the Carnegie Museum of Natural History. This was heavily and inaccurately restored with plaster using Allosaurus as a model before it was reverted to its original state.The first named fossil specimen which can be attributed to Tyrannosaurus rex consists of two partial vertebrae (one of which has been lost) found by Edward Drinker Cope in 1892. Cope believed that they belonged to an "agathaumid" (ceratopsid) dinosaur, and named them Manospondylus gigas, meaning "giant porous vertebra" in reverence to the numerous openings for blood vessels he found in the bone.[39] The M. gigas remains were later identified as those of a theropod rather than a ceratopsid, and H.F. Osborn recognized the similarity between M. gigas and Tyrannosaurus rex as early as 1917. However, due to the fragmentary nature of the Manospondylus vertebrae, Osborn did not synonymize the two genera.[40]
In June 2000, the Black Hills Institute located the type locality of M. gigas in South Dakota and unearthed more tyrannosaur bones there. These were judged to represent further remains of the same individual, and to be identical to those of Tyrannosaurus rex.[41] According to the rules of the International Code of Zoological Nomenclature (ICZN), the system that governs the scientific naming of animals, Manospondylus gigas should therefore have priority over Tyrannosaurus rex, because it was named first. However, the Fourth Edition of the ICZN, which took effect on 1 January 2000, states that "the prevailing usage must be maintained" when "the senior synonym or homonym has not been used as a valid name after 1899" and "the junior synonym or homonym has been used for a particular taxon, as its presumed valid name, in at least 25 works, published by at least 10 authors in the immediately preceding 50 years ..."[42] Tyrannosaurus rex may qualify as the valid name under these conditions and would most likely be considered a nomen protectum ("protected name") under the ICZN if it is ever formally published on, which it has not yet been. Manospondylus gigas could then be deemed a nomen oblitum ("forgotten name").[43]
Paleobiology
Life history
The identification of several specimens as juvenile Tyrannosaurus rex has allowed scientists to document ontogenetic changes in the species, estimate the lifespan, and determine how quickly the animals would have grown. The smallest known individual (LACM 28471, the "Jordan theropod") is estimated to have weighed only 30 kg (66 lb), while the largest, such as FMNH PR2081 ("Sue") most likely weighed over 5,400 kg (12,000 lb). Histologic analysis of Tyrannosaurus rex bones showed LACM 28471 had aged only 2 years when it died, while "Sue" was 28 years old, an age which may have been close to the maximum for the species.[4]
 A graph showing the hypothesized growth curve, body mass versus age (drawn in black, with other tyrannosaurids for comparison). Based on Erickson et al. 2004
A graph showing the hypothesized growth curve, body mass versus age (drawn in black, with other tyrannosaurids for comparison). Based on Erickson et al. 2004Histology has also allowed the age of other specimens to be determined. Growth curves can be developed when the ages of different specimens are plotted on a graph along with their mass. A Tyrannosaurus rex growth curve is S-shaped, with juveniles remaining under 1,800 kg (4,000 lb) until approximately 14 years of age, when body size began to increase dramatically. During this rapid growth phase, a young Tyrannosaurus rex would gain an average of 600 kg (1,300 lb) a year for the next four years. At 18 years of age, the curve plateaus again, indicating that growth slowed dramatically. For example, only 600 kg (1,300 lb) separated the 28-year-old "Sue" from a 22-year-old Canadian specimen (RTMP 81.12.1).[4] Another recent histological study performed by different workers corroborates these results, finding that rapid growth began to slow at around 16 years of age.[44] This sudden change in growth rate may indicate physical maturity, a hypothesis which is supported by the discovery of medullary tissue in the femur of a 16 to 20-year-old Tyrannosaurus rex from Montana (MOR 1125, also known as "B-rex"). Medullary tissue is found only in female birds during ovulation, indicating that "B-rex" was of reproductive age.[45] Further study indicates an age of 18 for this specimen.[46] Other tyrannosaurids exhibit extremely similar growth curves, although with lower growth rates corresponding to their lower adult sizes.[47]
Over half of the known Tyrannosaurus rex specimens appear to have died within six years of reaching sexual maturity, a pattern which is also seen in other tyrannosaurs and in some large, long-lived birds and mammals today. These species are characterized by high infant mortality rates, followed by relatively low mortality among juveniles. Mortality increases again following sexual maturity, partly due to the stresses of reproduction. One study suggests that the rarity of juvenile Tyrannosaurus rex fossils is due in part to low juvenile mortality rates; the animals were not dying in large numbers at these ages, and so were not often fossilized. However, this rarity may also be due to the incompleteness of the fossil record or to the bias of fossil collectors towards larger, more spectacular specimens.[47]
Sexual dimorphism
As the number of specimens increased, scientists began to analyze the variation between individuals and discovered what appeared to be two distinct body types, or morphs, similar to some other theropod species. As one of these morphs was more solidly built, it was termed the 'robust' morph while the other was termed 'gracile'. Several morphological differences associated with the two morphs were used to analyze sexual dimorphism in Tyrannosaurus rex, with the 'robust' morph usually suggested to be female. For example, the pelvis of several 'robust' specimens seemed to be wider, perhaps to allow the passage of eggs.[48] It was also thought that the 'robust' morphology correlated with a reduced chevron on the first tail vertebra, also ostensibly to allow eggs to pass out of the reproductive tract, as had been erroneously reported for crocodiles.[49]
 Skeleton casts mounted in a mating position, Jurassic Museum of Asturias
Skeleton casts mounted in a mating position, Jurassic Museum of AsturiasIn recent years, evidence for sexual dimorphism has been weakened. A 2005 study reported that previous claims of sexual dimorphism in crocodile chevron anatomy were in error, casting doubt on the existence of similar dimorphism between Tyrannosaurus rex genders.[50] A full-sized chevron was discovered on the first tail vertebra of "Sue", an extremely robust individual, indicating that this feature could not be used to differentiate the two morphs anyway. As Tyrannosaurus rex specimens have been found from Saskatchewan to New Mexico, differences between individuals may be indicative of geographic variation rather than sexual dimorphism. The differences could also be age-related, with 'robust' individuals being older animals.[14]
Only a single Tyrannosaurus rex specimen has been conclusively shown to belong to a specific gender. Examination of "B-rex" demonstrated the preservation of soft tissue within several bones. Some of this tissue has been identified as a medullary tissue, a specialized tissue grown only in modern birds as a source of calcium for the production of eggshell during ovulation. As only female birds lay eggs, medullary tissue is only found naturally in females, although males are capable of producing it when injected with female reproductive hormones like estrogen. This strongly suggests that "B-rex" was female, and that she died during ovulation.[45] Recent research has shown that medullary tissue is never found in crocodiles, which are thought to be the closest living relatives of dinosaurs, aside from birds. The shared presence of medullary tissue in birds and theropod dinosaurs is further evidence of the close evolutionary relationship between the two.[51]
Posture
 Outdated reconstruction (by Charles R. Knight), showing upright pose
Outdated reconstruction (by Charles R. Knight), showing upright poseLike many bipedal dinosaurs, Tyrannosaurus rex was historically depicted as a 'living tripod', with the body at 45 degrees or less from the vertical and the tail dragging along the ground, similar to a kangaroo. This concept dates from Joseph Leidy's 1865 reconstruction of Hadrosaurus, the first to depict a dinosaur in a bipedal posture.[52] Henry Fairfield Osborn, former president of the American Museum of Natural History (AMNH) in New York City, who believed the creature stood upright, further reinforced the notion after unveiling the first complete Tyrannosaurus rex skeleton in 1915. It stood in this upright pose for 77 years, until it was dismantled in 1992.[53] By 1970, scientists realized this pose was incorrect and could not have been maintained by a living animal, as it would have resulted in the dislocation or weakening of several joints, including the hips and the articulation between the head and the spinal column.[54] The inaccurate AMNH mount inspired similar depictions in many films and paintings (such as Rudolph Zallinger's famous mural The Age Of Reptiles in Yale University's Peabody Museum of Natural History)[55] until the 1990s, when films such as Jurassic Park introduced a more accurate posture to the general public. Modern representations in museums, art, and film show Tyrannosaurus rex with its body approximately parallel to the ground and tail extended behind the body to balance the head.[23]
Arms
 The forelimbs might have been used to help T. rex rise from a resting pose, as seen here
The forelimbs might have been used to help T. rex rise from a resting pose, as seen hereWhen Tyrannosaurus rex was first discovered, the humerus was the only element of the forelimb known.[56] For the initial mounted skeleton as seen by the public in 1915, Osborn substituted longer, three-fingered forelimbs like those of Allosaurus.[40] However, a year earlier, Lawrence Lambe described the short, two-fingered forelimbs of the closely related Gorgosaurus.[57] This strongly suggested that Tyrannosaurus rex had similar forelimbs, but this hypothesis was not confirmed until the first complete Tyrannosaurus rex forelimbs were identified in 1989, belonging to MOR 555 (the "Wankel rex").[58] The remains of "Sue" also include complete forelimbs.[14] Tyrannosaurus rex arms are very small relative to overall body size, measuring only 1 metre (3.3 ft) long. However, they are not vestigial but instead show large areas for muscle attachment, indicating considerable strength. This was recognized as early as 1906 by Osborn, who speculated that the forelimbs may have been used to grasp a mate during copulation.[59] It has also been suggested that the forelimbs were used to assist the animal in rising from a prone position.[54] Another possibility is that the forelimbs held struggling prey while it was killed by the tyrannosaur's enormous jaws. This hypothesis may be supported by biomechanical analysis.
 Diagram illustrating arm anatomy
Diagram illustrating arm anatomyTyrannosaurus rex forelimb bones exhibit extremely thick cortical bone, indicating that they were developed to withstand heavy loads. The biceps brachii muscle of a full-grown Tyrannosaurus rex was capable of lifting 199 kilograms (439 lb) by itself; other muscles such as the brachialis would work along with the biceps to make elbow flexion even more powerful. The M. biceps muscle of T. rex was 3.5 times as powerful as the human equivalent. A Tyrannosaurus rex forearm also had a reduced range of motion, with the shoulder and elbow joints allowing only 40 and 45 degrees of motion, respectively. In contrast, the same two joints in Deinonychus allow up to 88 and 130 degrees of motion, respectively, while a human arm can rotate 360 degrees at the shoulder and move through 165 degrees at the elbow. The heavy build of the arm bones, extreme strength of the muscles, and limited range of motion may indicate a system evolved to hold fast despite the stresses of a struggling prey animal. Carpenter and Smith dismissed notions that the forelimbs were useless or that Tyrannosaurus rex was an obligate scavenger.[60]
Legs

The metatarsal of the middle main toe (toe III, see this image) tapers to nothing.[61]
Soft tissue
In the March 2005 issue of Science, Mary Higby Schweitzer of North Carolina State University and colleagues announced the recovery of soft tissue from the marrow cavity of a fossilized leg bone, from a Tyrannosaurus rex. The bone had been intentionally, though reluctantly, broken for shipping and then not preserved in the normal manner, specifically because Schweitzer was hoping to test it for soft tissue.[62] Designated as the Museum of the Rockies specimen 1125, or MOR 1125, the dinosaur was previously excavated from the Hell Creek Formation. Flexible, bifurcating blood vessels and fibrous but elastic bone matrix tissue were recognized. In addition, microstructures resembling blood cells were found inside the matrix and vessels. The structures bear resemblance to ostrich blood cells and vessels. Whether an unknown process, distinct from normal fossilization, preserved the material, or the material is original, the researchers do not know, and they are careful not to make any claims about preservation.[63] If it is found to be original material, any surviving proteins may be used as a means of indirectly guessing some of the DNA content of the dinosaurs involved, because each protein is typically created by a specific gene. The absence of previous finds may merely be the result of people assuming preserved tissue was impossible, therefore simply not looking. Since the first, two more tyrannosaurs and a hadrosaur have also been found to have such tissue-like structures.[62] Research on some of the tissues involved has suggested that birds are closer relatives to tyrannosaurs than other modern animals.[64]
 T. rex femur (MOR 1125) from which demineralized matrix and peptides (insets) were obtained
T. rex femur (MOR 1125) from which demineralized matrix and peptides (insets) were obtainedIn studies reported in the journal Science in April 2007, Asara and colleagues concluded that seven traces of collagen proteins detected in purified Tyrannosaurus rex bone most closely match those reported in chickens, followed by frogs and newts. The discovery of proteins from a creature tens of millions of years old, along with similar traces the team found in a mastodon bone at least 160,000 years old, upends the conventional view of fossils and may shift paleontologists' focus from bone hunting to biochemistry. Until these finds, most scientists presumed that fossilization replaced all living tissue with inert minerals. Paleontologist Hans Larsson of McGill University in Montreal, who was not part of the studies, called the finds "a milestone", and suggested that dinosaurs could "enter the field of molecular biology and really slingshot paleontology into the modern world".[65]
Subsequent studies in April 2008 confirmed the close connection of Tyrannosaurus rex to modern birds. Postdoctoral biology researcher Chris Organ at Harvard University announced, "With more data, they would probably be able to place T. rex on the evolutionary tree between alligators and chickens and ostriches." Co-author John M. Asara added, "We also show that it groups better with birds than modern reptiles, such as alligators and green anole lizards."[66]
The presumed soft tissue was called into question by Thomas Kaye of the University of Washington and his co-authors in 2008. They contend that what was really inside the tyrannosaur bone was slimy biofilm created by bacteria that coated the voids once occupied by blood vessels and cells.[67] The researchers found that what previously had been identified as remnants of blood cells, because of the presence of iron, were actually framboids, microscopic mineral spheres bearing iron. They found similar spheres in a variety of other fossils from various periods, including an ammonite. In the ammonite they found the spheres in a place where the iron they contain could not have had any relationship to the presence of blood.[68] However, Schweitzer has strongly criticized Kaye's claims and maintains that she really did find blood cells, and argues that there’s no reported evidence that biofilms can produce branching, hollow tubes like those noted in her study.[69] San Antonio, Schweitzer and colleagues published an analysis in 2011 of what parts of the collagen had been recovered, finding that it was the inner parts of the collagen coil that had been preserved, as would have been expected from a long period of protein degradation.[70]
Skin and feathers
Main article: Feathered dinosaurs Restoration of a young Tyrannosaurus, depicted with filamentous feathers.
Restoration of a young Tyrannosaurus, depicted with filamentous feathers.In 2004, the scientific journal Nature published a report describing an early tyrannosauroid, Dilong paradoxus, from the famous Yixian Formation of China. As with many other theropods discovered in the Yixian, the fossil skeleton was preserved with a coat of filamentous structures which are commonly recognized as the precursors of feathers. It has also been proposed that Tyrannosaurus and other closely related tyrannosaurids had such protofeathers. However, skin impressions from large tyrannosaurid specimens show mosaic scales.[71] While it is possible that protofeathers existed on parts of the body which have not been preserved, a lack of insulatory body covering is consistent with modern multi-ton mammals such as elephants, hippopotamus, and most species of rhinoceros. As an object increases in size, its ability to retain heat increases due to its decreasing surface area-to-volume ratio. Therefore, as large animals evolve in or disperse into warm climates, a coat of fur or feathers loses its selective advantage for thermal insulation and can instead become a disadvantage, as the insulation traps excess heat inside the body, possibly overheating the animal. Protofeathers may also have been secondarily lost during the evolution of large tyrannosaurids like Tyrannosaurus, especially in warm Cretaceous climates.[72]
Thermoregulation
Main article: Physiology of dinosaursTyrannosaurus, like most dinosaurs, was long thought to have an ectothermic ("cold-blooded") reptilian metabolism. The idea of dinosaur ectothermy was challenged by scientists like Robert T. Bakker and John Ostrom in the early years of the "Dinosaur Renaissance", beginning in the late 1960s.[73][74] Tyrannosaurus rex itself was claimed to have been endothermic ("warm-blooded"), implying a very active lifestyle.[6] Since then, several paleontologists have sought to determine the ability of Tyrannosaurus to regulate its body temperature. Histological evidence of high growth rates in young Tyrannosaurus rex, comparable to those of mammals and birds, may support the hypothesis of a high metabolism. Growth curves indicate that, as in mammals and birds, Tyrannosaurus rex growth was limited mostly to immature animals, rather than the indeterminate growth seen in most other vertebrates.[44]
Oxygen isotope ratios in fossilized bone are sometimes used to determine the temperature at which the bone was deposited, as the ratio between certain isotopes correlates with temperature. In one specimen, the isotope ratios in bones from different parts of the body indicated a temperature difference of no more than 4 to 5 °C (7 to 9 °F) between the vertebrae of the torso and the tibia of the lower leg. This small temperature range between the body core and the extremities was claimed by paleontologist Reese Barrick and geochemist William Showers to indicate that Tyrannosaurus rex maintained a constant internal body temperature (homeothermy) and that it enjoyed a metabolism somewhere between ectothermic reptiles and endothermic mammals.[75] Other scientists have pointed out that the ratio of oxygen isotopes in the fossils today does not necessarily represent the same ratio in the distant past, and may have been altered during or after fossilization (diagenesis).[76] Barrick and Showers have defended their conclusions in subsequent papers, finding similar results in another theropod dinosaur from a different continent and tens of millions of years earlier in time (Giganotosaurus).[77] Ornithischian dinosaurs also showed evidence of homeothermy, while varanid lizards from the same formation did not.[78] Even if Tyrannosaurus rex does exhibit evidence of homeothermy, it does not necessarily mean that it was endothermic. Such thermoregulation may also be explained by gigantothermy, as in some living sea turtles.[79][80]
Footprints
 Probable footprint from New Mexico
Probable footprint from New MexicoTwo isolated fossilized footprints have been tentatively assigned to Tyrannosaurus rex. The first was discovered at Philmont Scout Ranch, New Mexico, in 1983 by American geologist Charles Pillmore. Originally thought to belong to a hadrosaurid, examination of the footprint revealed a large 'heel' unknown in ornithopod dinosaur tracks, and traces of what may have been a hallux, the dewclaw-like fourth digit of the tyrannosaur foot. The footprint was published as the ichnogenus Tyrannosauripus pillmorei in 1994, by Martin Lockley and Adrian Hunt. Lockley and Hunt suggested that it was very likely the track was made by a Tyrannosaurus rex, which would make it the first known footprint from this species. The track was made in what was once a vegetated wetland mud flat. It measures 83 centimetres (33 in) long by 71 centimetres (28 in) wide.[81]
A second footprint that may have been made by a Tyrannosaurus was first reported in 2007 by British paleontologist Phil Manning, from the Hell Creek Formation of Montana. This second track measures 76 centimetres (30 in) long, shorter than the track described by Lockley and Hunt. Whether or not the track was made by Tyrannosaurus is unclear, though Tyrannosaurus and Nanotyrannus are the only large theropods known to have existed in the Hell Creek Formation. Further study of the track (a full description has not yet been published) will compare the Montana track with the one found in New Mexico.[82]
Locomotion
There are two main issues concerning the locomotory abilities of Tyrannosaurus: how well it could turn; and what its maximum straight-line speed was likely to have been. Both are relevant to the debate about whether it was a hunter or a scavenger (see below).
Tyrannosaurus may have been slow to turn, possibly taking one to two seconds to turn only 45° — an amount that humans, being vertically oriented and tailless, can spin in a fraction of a second.[83] The cause of the difficulty is rotational inertia, since much of Tyrannosaurus’ mass was some distance from its center of gravity, like a human carrying a heavy timber — although it might have reduced the average distance by arching its back and tail and pulling its head and forelimbs close to its body, rather like the way ice skaters pull their arms closer in order to spin faster.[84]
 Replica of a sequence of theropod footprints attributed to Megalosaurus at OUMNH. No such sequence has yet been reported for tyrannosaurs, making gait and speed estimates difficult
Replica of a sequence of theropod footprints attributed to Megalosaurus at OUMNH. No such sequence has yet been reported for tyrannosaurs, making gait and speed estimates difficultScientists have produced a wide range of maximum speed estimates, mostly around 11 metres per second (40 km/h; 25 mph), but a few as low as 5–11 metres per second (18–40 km/h; 11–25 mph), and a few as high as 20 metres per second (72 km/h; 45 mph). Researchers have to rely on various estimating techniques because, while there are many tracks of very large theropods walking, so far none have been found of very large theropods running—and this absence may indicate that they did not run.[85] Scientists who think that Tyrannosaurus was able to run point out that hollow bones and other features that would have lightened its body may have kept adult weight to a mere 4.5 metric tons (5.0 short tons) or so, or that other animals like ostriches and horses with long, flexible legs are able to achieve high speeds through slower but longer strides. Additionally, some have argued that Tyrannosaurus had relatively larger leg muscles than any animal alive today, which could have enabled fast running 40–70 kilometres per hour (25–43 mph).[86]
Jack Horner and Don Lessem argued in 1993 that Tyrannosaurus was slow and probably could not run (no airborne phase in mid-stride), because its ratio of femur (thigh bone) to tibia (shin bone) length was greater than 1, as in most large theropods and like a modern elephant.[58] However, Holtz (1998) noted that tyrannosaurids and some closely related groups had significantly longer distal hindlimb components (shin plus foot plus toes) relative to the femur length than most other theropods), and that tyrannosaurids and their close relatives had a tightly interlocked metatarsus that more effectively transmitted locomotory forces from the foot to the lower leg than in earlier theropods ("metatarsus" means the foot bones, which function as part of the leg in digitigrade animals). He therefore concluded that tyrannosaurids and their close relatives were the fastest large theropods.[87]
Skeletal anatomy of a T. rex right legChristiansen (1998) estimated that the leg bones of Tyrannosaurus were not significantly stronger than those of elephants, which are relatively limited in their top speed and never actually run (there is no airborne phase), and hence proposed that the dinosaur's maximum speed would have been about 11 metres per second (40 km/h; 25 mph), which is about the speed of a human sprinter. But he also noted that such estimates depend on many dubious assumptions.[88]
Farlow and colleagues (1995) have argued that a Tyrannosaurus weighing 5.4 metric tons (6.0 short tons) to 7.3 metric tons (8.0 short tons) would have been critically or even fatally injured if it had fallen while moving quickly, since its torso would have slammed into the ground at a deceleration of 6 g (six times the acceleration due to gravity, or about 60 meters/s²) and its tiny arms could not have reduced the impact.[7] However, giraffes have been known to gallop at 50 kilometres per hour (31 mph), despite the risk that they might break a leg or worse, which can be fatal even in a "safe" environment such as a zoo.[89][90] Thus it is quite possible that Tyrannosaurus also moved fast when necessary and had to accept such risks.[91][92]
Most recent research on Tyrannosaurus locomotion does not support speeds faster than 40 kilometres per hour (25 mph), i.e. moderate-speed running. For example, a 2002 paper in the journal Nature used a mathematical model (validated by applying it to three living animals, alligators, chickens, and humans; additionally later eight more species including emus and ostriches[85]) to gauge the leg muscle mass needed for fast running (over 40 km/h or 25 mph).[86] They found that proposed top speeds in excess of 40 kilometres per hour (25 mph) were unfeasible, because they would require very large leg muscles (more than approximately 40–86% of total body mass). Even moderately fast speeds would have required large leg muscles. This discussion is difficult to resolve, as it is unknown how large the leg muscles actually were in Tyrannosaurus. If they were smaller, only 18 kilometres per hour (11 mph) walking/jogging might have been possible.[86]
 Restoration of T. rex in walking posture
Restoration of T. rex in walking postureA study in 2007 used computer models to estimate running speeds, based on data taken directly from fossils, and claimed that Tyrannosaurus rex had a top running speed of 8 metres per second (29 km/h; 18 mph). An average professional football (soccer) player would be slightly slower, while a human sprinter can reach 12 metres per second (43 km/h; 27 mph). Note that these computer models predict a top speed of 17.8 metres per second (64 km/h; 40 mph) for a 3-kilogram (6.6 lb) Compsognathus[93][94] (probably a juvenile individual).[95]
Those who argue that Tyrannosaurus was incapable of running estimate the top speed of Tyrannosaurus at about 17 kilometres per hour (11 mph). This is still faster than its most likely prey species, hadrosaurids and ceratopsians.[86] In addition, some advocates of the idea that Tyrannosaurus was a predator claim that tyrannosaur running speed is not important, since it may have been slow but still faster than its probable prey.[96] However, Paul and Christiansen (2000) argued that at least the later ceratopsians had upright forelimbs and the larger species may have been as fast as rhinos.[97] Healed Tyrannosaurus bite wounds on ceratopsian fossils are interpreted as evidence of attacks on living ceratopsians (see below). If the ceratopsians that lived alongside Tyrannosaurus were fast, that casts doubt on the argument that Tyrannosaurus did not have to be fast to catch its prey.[92]
Feeding strategies
The debate about whether Tyrannosaurus was a predator or a pure scavenger is as old as the debate about its locomotion. Lambe (1917) described a good skeleton of Tyrannosaurus’ close relative Gorgosaurus and concluded that it and therefore also Tyrannosaurus was a pure scavenger, because the Gorgosaurus’ teeth showed hardly any wear.[98] This argument is no longer taken seriously, because theropods replaced their teeth quite rapidly. Ever since the first discovery of Tyrannosaurus most scientists have speculated that it was a predator; like modern large predators it would readily scavenge or steal another predator's kill if it had the opportunity.[99]
Paleontologist Jack Horner has been a major advocate of the idea that Tyrannosaurus was exclusively a scavenger and did not engage in active hunting at all,[58][100][101] though Horner himself has claimed that he never published this idea in the peer reviewed scientific literature and used it mainly as a tool to teach a popular audience, particularly children, the dangers of making assumptions in science (such as assuming T. rex was a hunter) without using evidence.[102] Nevertheless, Horner presented several arguments in the popular literature to support the pure scavenger hypothesis:
 Cast of the braincase at the Australian Museum, Sydney
Cast of the braincase at the Australian Museum, Sydney- Tyrannosaur arms are short when compared to other known predators. Horner argues that the arms were too short to make the necessary gripping force to hold on to prey.[103]
- Tyrannosaurs had large olfactory bulbs and olfactory nerves (relative to their brain size). These suggest a highly developed sense of smell which could sniff out carcasses over great distances, as modern vultures do. Research on the olfactory bulbs of dinosaurs has shown that Tyrannosaurus had the most highly developed sense of smell of 21 sampled dinosaurs.[104] Opponents of the pure scavenger hypothesis have used the example of vultures in the opposite way, arguing that the scavenger hypothesis is implausible because the only modern pure scavengers are large gliding birds, which use their keen senses and energy-efficient gliding to cover vast areas economically.[105] However, researchers from Glasgow concluded that an ecosystem as productive as the current Serengeti would provide sufficient carrion for a large theropod scavenger, although the theropod might have had to be cold-blooded in order to get more calories from carrion than it spent on foraging (see Metabolism of dinosaurs). They also suggested that modern ecosystems like Serengeti have no large terrestrial scavengers because gliding birds now do the job much more efficiently, while large theropods did not face competition for the scavenger ecological niche from gliding birds.[106]
- Tyrannosaur teeth could crush bone, and therefore could extract as much food (bone marrow) as possible from carcass remnants, usually the least nutritious parts. Karen Chin and colleagues have found bone fragments in coprolites (fossilized feces) that they attribute to tyrannosaurs, but point out that a tyrannosaur's teeth were not well adapted to systematically chewing bone like hyenas do to extract marrow.[107]
- Since at least some of Tyrannosaurus's potential prey could move quickly, evidence that it walked instead of ran could indicate that it was a scavenger.[100][108] On the other hand, recent analyses suggest that Tyrannosaurus, while slower than large modern terrestrial predators, may well have been fast enough to prey on large hadrosaurs and ceratopsians.[86][96]
 The eye-sockets faced mainly forwards, giving it good binocular vision
The eye-sockets faced mainly forwards, giving it good binocular visionOther evidence suggests hunting behavior in Tyrannosaurus. The eye-sockets of tyrannosaurs are positioned so that the eyes would point forward, giving them binocular vision slightly better than that of modern hawks. Horner also pointed out that the tyrannosaur lineage had a history of steadily improving binocular vision. It is not obvious why natural selection would have favored this long-term trend if tyrannosaurs had been pure scavengers, which would not have needed the advanced depth perception that stereoscopic vision provides.[17][18] In modern animals, binocular vision is found mainly in predators.
 Restoration (based on MOR 980) with parasite infections, which might be the cause of scars seen in the skulls of several specimens that were previously explained by intraspecific attacks[109]
Restoration (based on MOR 980) with parasite infections, which might be the cause of scars seen in the skulls of several specimens that were previously explained by intraspecific attacks[109]A skeleton of the hadrosaurid Edmontosaurus annectens has been described from Montana with healed tyrannosaur-inflicted damage on its tail vertebrae. The fact that the damage seems to have healed suggests that the Edmontosaurus survived a tyrannosaur's attack on a living target, i.e. the tyrannosaur had attempted active predation.[110] There is also evidence for an aggressive interaction between a Triceratops and a Tyrannosaurus in the form of partially healed tyrannosaur tooth marks on a Triceratops brow horn and squamosal (a bone of the neck frill); the bitten horn is also broken, with new bone growth after the break. It is not known what the exact nature of the interaction was, though: either animal could have been the aggressor.[111] When examining Sue, paleontologist Pete Larson found a broken and healed fibula and tail vertebrae, scarred facial bones and a tooth from another Tyrannosaurus embedded in a neck vertebra. If correct, these might be strong evidence for aggressive behavior between tyrannosaurs but whether it would have been competition for food and mates or active cannibalism is unclear.[112] However, further recent investigation of these purported wounds has shown that most are infections rather than injuries (or simply damage to the fossil after death) and the few injuries are too general to be indicative of intraspecific conflict.[100] A 2009 study showed that holes in the skulls of several specimens might have been caused by Trichomonas-like parasites that commonly infect avians.[109]
Some researchers argue that if Tyrannosaurus were a scavenger, another dinosaur had to be the top predator in the Amerasian Upper Cretaceous. Top prey was the larger marginocephalians and ornithopods. The other tyrannosaurids share so many characteristics that only small dromaeosaurs and troodontids remain as feasible top predators. In this light, scavenger hypothesis adherents have suggested that the size and power of tyrannosaurs allowed them to steal kills from smaller predators,[108] although they may have had a hard time finding enough meat to scavenge, being outnumbered by smaller theropods.[113] Most paleontologists accept that Tyrannosaurus was both an active predator and a scavenger like most large carnivores.
Tyrannosaurus may have had infectious saliva used to kill its prey. This theory was first proposed by William Abler.[114] Abler examined the teeth of tyrannosaurids between each tooth serration; the serrations may have held pieces of carcass with bacteria, giving Tyrannosaurus a deadly, infectious bite much like the Komodo Dragon was thought to have. However, Jack Horner regards Tyrannosaurus tooth serrations as more like cubes in shape than the serrations on a Komodo monitor's teeth, which are rounded.[115] All forms of saliva contain possibly hazardous bacteria, so the prospect of it being used as a method of predation is disputable.
Cannibalism
A study from Currie, Horner, Erickson and Longrich in 2010 has been put forward as evidence of cannibalism in the genus Tyrannosaurus.[116] They studied some Tyrannosaurus specimens with tooth marks in the bones, attributable to the same genus. The tooth marks were identified in the humerus, foot bones and metatarsals, and this was seen as evidence for opportunistic scavenging, rather than wounds caused by intraspecific combat. In a fight, they proposed it would be difficult to reach down to bite in the feet of a rival, making it more likely that the bitemarks were made in a carcass. As the bitemarks were made in body parts with relatively scantly amounts of flesh, it is suggested that the Tyrannosaurus was feeding on a cadaver in which the more fleshy parts already had been eaten up.[116] They were also open to the possibility that other tyrannosaurids practiced cannibalism.
Paleopathology
In 2001, Bruce Rothschild and others published a study examining evidence for stress fractures and tendon avulsions in theropod dinosaurs and the implications for their behavior. Since stress fractures are caused by repeated trauma rather than singular events they are more likely to be caused by regular behavior than other types of injuries. Of the 81 Tyrannosaurus foot bones examined in the study one was found to have a stress fracture, while none of the 10 hand bones were found to have stress fractures. The researchers found tendon avulsions only among Tyrannosaurus and Allosaurus. An avulsion injury left a divot on the humerus of Sue the T. rex, apparently located at the origin of the deltoid or teres major muscles. The presence of avulsion injuries being limited to a the forelimb and shoulder in both Tyrannosaurus and Allosaurus suggests that theropods may have had a musculature more complex and functionally different than those of birds. The researchers concluded that Sue's tendon avulsion was probably obtained from struggling prey. The presence of stress fractures and tendon avulsions in general provide evidence for a "very active" predation-based diet rather than obligate scavenging.[117]
History
 Skeletal restoration by William D. Matthew from 1905, the first reconstruction of this dinosaur ever published[118]
Skeletal restoration by William D. Matthew from 1905, the first reconstruction of this dinosaur ever published[118]Henry Fairfield Osborn, president of the American Museum of Natural History, named Tyrannosaurus rex in 1905. The generic name is derived from the Greek words τυράννος (tyrannos, meaning "tyrant") and σαύρος (sauros, meaning "lizard"). Osborn used the Latin word rex, meaning "king", for the specific name. The full binomial therefore translates to "tyrant lizard the king" or "King Tyrant Lizard",[119] emphasizing the animal's size and perceived dominance over other species of the time.[56]
Earliest finds
Teeth from what is now documented as a Tyrannosaurus rex were found in 1874 by A. Lakes near Golden, Colorado. In the early 1890s, J. B. Hatcher collected postcranial elements in eastern Wyoming. The fossils were believed to be from a large species of Ornithomimus (O. grandis) but are now considered Tyrannosaurus rex. Vertebral fragments found by E. D. Cope in western South Dakota in 1892 and named as Manospondylus gigas have also been recognized as belonging to Tyrannosaurus rex.[39]
 Scale model of the never-completed exhibit planned for the American Museum of Natural History by H.F. Osborn
Scale model of the never-completed exhibit planned for the American Museum of Natural History by H.F. OsbornBarnum Brown, assistant curator of the American Museum of Natural History, found the first partial skeleton of Tyrannosaurus rex in eastern Wyoming in 1900. H. F. Osborn originally named this skeleton Dynamosaurus imperiosus in a paper in 1905. Brown found another partial skeleton in the Hell Creek Formation in Montana in 1902. Osborn used this holotype to describe Tyrannosaurus rex in the same paper in which D. imperiosus was described.[56] In 1906, Osborn recognized the two as synonyms, and acted as first revisor by selecting Tyrannosaurus as the valid name.[59] The original Dynamosaurus material resides in the collections of the Natural History Museum, London.[120]
In total, Brown found five Tyrannosaurus partial skeletons. In 1941, Brown's 1902 find was sold to the Carnegie Museum of Natural History in Pittsburgh, Pennsylvania. Brown's fourth and largest find, also from Hell Creek, is on display in the American Museum of Natural History in New York.[58]
Although there are numerous skeletons in the world, only one track has been documented — at Philmont Scout Ranch in northeast New Mexico. It was discovered in 1983 and identified and documented in 1994.[121]
Notable specimens
Main article: Specimens of Tyrannosaurus "Sue" specimen, Field Museum of Natural History, Chicago
"Sue" specimen, Field Museum of Natural History, ChicagoSue Hendrickson, amateur paleontologist, discovered the most complete (approximately 85%) and, until 2001, the largest, Tyrannosaurus fossil skeleton known in the Hell Creek Formation near Faith, South Dakota, on 12 August 1990. This Tyrannosaurus, nicknamed "Sue" in her honor, was the object of a legal battle over its ownership. In 1997 this was settled in favor of Maurice Williams, the original land owner. The fossil collection was purchased by the Field Museum of Natural History at auction for USD 7.6 million, making it the most expensive dinosaur skeleton to date. From 1998 to 1999 Field Museum of Natural History preparators spent over 25,000 man-hours taking the rock off each of the bones.[122] The bones were then shipped off to New Jersey where the mount was made. The finished mount was then taken apart, and along with the bones, shipped back to Chicago for the final assembly. The mounted skeleton opened to the public on May 17, 2000 in the great hall (Stanley Field Hall) at the Field Museum of Natural History. A study of this specimen's fossilized bones showed that "Sue" reached full size at age 19 and died at age 28, the longest any tyrannosaur is known to have lived.[123] Early speculation that Sue may have died from a bite to the back of the head was not confirmed. Though subsequent study showed many pathologies in the skeleton, no bite marks were found.[124] Damage to the back of the skull may have been caused by post-mortem trampling. Recent speculation indicates that "Sue" may have died of starvation after contracting a parasitic infection from eating diseased meat; the resulting infection would have caused inflammation in the throat, ultimately leading "Sue" to starve because she could no longer swallow food. This hypothesis is substantiated by smooth-edged holes in her skull which are similar to those caused in modern-day birds that contract the same parasite.[125]
Another Tyrannosaurus, nicknamed "Stan", in honor of amateur paleontologist Stan Sacrison, was found in the Hell Creek Formation near Buffalo, South Dakota, in the spring of 1987. It was not collected until 1992, as it was mistakenly thought to be a Triceratops skeleton. Stan is 63% complete and is on display in the Black Hills Institute of Geological Research in Hill City, South Dakota, after an extensive world tour during 1995 and 1996.[126] This tyrannosaur, too, was found to have many bone pathologies, including broken and healed ribs, a broken (and healed) neck and a spectacular hole in the back of its head, about the size of a Tyrannosaurus tooth.[127]
 Size of various specimens compared with a human
Size of various specimens compared with a humanIn the summer of 2000, Jack Horner discovered five Tyrannosaurus skeletons near the Fort Peck Reservoir in Montana. One of the specimens, dubbed "C. rex", was reported to be perhaps the largest Tyrannosaurus ever found.[128]
In 2001, a 50% complete skeleton of a juvenile Tyrannosaurus was discovered in the Hell Creek Formation in Montana, by a crew from the Burpee Museum of Natural History of Rockford, Illinois. Dubbed "Jane", the find was initially considered the first known skeleton of the pygmy tyrannosaurid Nanotyrannus but subsequent research has revealed that it is more likely a juvenile Tyrannosaurus.[129] It is the most complete and best preserved juvenile example known to date. Jane has been examined by Jack Horner, Pete Larson, Robert Bakker, Greg Erickson, and several other renowned paleontologists, because of the uniqueness of her age. "Jane" is currently on exhibit at the Burpee Museum of Natural History in Rockford, Illinois.[130][131]
In a press release on 7 April 2006, Montana State University revealed that it possessed the largest Tyrannosaurus skull yet discovered. Discovered in the 1960s and only recently reconstructed, the skull measures 59 inches (150 cm) long compared to the 55.4 inches (141 cm) of "Sue's" skull, a difference of 6.5%.[132][133]
Appearances in popular culture
Main article: Tyrannosaurus in popular cultureSince it was first described in 1905, Tyrannosaurus rex has become the most widely recognized dinosaur species in popular culture. It is the only dinosaur that is commonly known to the general public by its full scientific name (binomial name) (Tyrannosaurus rex), and the scientific abbreviation T. rex has also come into wide usage.[14] Robert T. Bakker notes this in The Dinosaur Heresies and explains that a name like "Tyrannosaurus rex is just irresistible to the tongue."[6]
References
- ^ Hicks, J.F., Johnson, K.R., Obradovich, J.D., Tauxe, L. and Clark, D. (2002). "Magnetostratigraphy and geochronology of the Hell Creek and basal Fort Union Formations of southwestern North Dakota and a recalibration of the Cretaceous–Tertiary Boundary", in J.H. Hartman, K.R. Johnson & D.J. Nichols (eds.), The Hell Creek Formation and the Cretaceous–Tertiary boundary in the northern Great Plains: An integrated continental record of the end of the Cretaceous. GSA Special Paper, 361: 35–55.
- ^ a b Henderson DM (January 1, 1999). "Estimating the masses and centers of mass of extinct animals by 3-D mathematical slicing". Paleobiology 25 (1): 88–106. http://paleobiol.geoscienceworld.org/cgi/content/abstract/25/1/88.
- ^ a b c "Sue's vital statistics". Sue at the Field Museum. Field Museum of Natural History. http://www.fieldmuseum.org/sue/about_vital.asp. Retrieved 2007-09-15.
- ^ a b c d Erickson, Gregory M., GM; Makovicky, Peter J.; Currie, Philip J.; Norell, Mark A.; Yerby, Scott A.; & Brochu, Christopher A. (2004). "Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs". Nature 430 (7001): 772–775. doi:10.1038/nature02699. PMID 15306807.
- ^ Anderson, JF; Hall-Martin AJ Russell DA (1985). "Long bone circumference and weight in mammals, birds and dinosaurs". Journal of Zoology 207 (1): 53–61. doi:10.1111/j.1469-7998.1985.tb04915.x.
- ^ a b c Bakker, Robert T. (1986). The Dinosaur Heresies. New York: Kensington Publishing. ISBN 0-688-04287-2. OCLC 13699558.[page needed]
- ^ a b Farlow, JO; Smith MB, Robinson JM (1995). "Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex". Journal of Vertebrate Paleontology 15 (4): 713–725. doi:10.1080/02724634.1995.10011257. http://www.vertpaleo.org/publications/jvp/15-713-725.cfm.
- ^ Seebacher, Frank. (2001). "A new method to calculate allometric length–mass relationships of dinosaurs". Journal of Vertebrate Paleontology 21 (1): 51–60. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2.
- ^ Christiansen, Per; & Fariña, Richard A. (2004). "Mass prediction in theropod dinosaurs". Historical Biology 16 (2–4): 85–92. doi:10.1080/08912960412331284313.
- ^ Boardman T.J., Packard G.C., Birchard G.F. (2009). "Allometric equations for predicting body mass of dinosaurs". Journal of Zoology 279 (1): 102–110. doi:10.1111/j.1469-7998.2009.00594.x.
- ^ Hutchinson J.R., Bates K.T., Molnar J., Allen V & Makovicky P.J. (2011), "A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth", PLoS ONE 6(10): e26037. doi:10.1371/journal.pone.0026037
- ^ dal Sasso, Cristiano; Maganuco, Simone; Buffetaut, Eric; & Mendez, Marcos A. (2005). "New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities". Journal of Vertebrate Paleontology 25 (4): 888–896. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1671%2F0272-4634%282005%29025%5B0888%3ANIOTSO%5D2.0.CO%3B2.
- ^ Calvo, Jorge O.; Rodolfo Coria (December 1998). "New specimen of Giganotosaurus carolinii (Coria & Salgado, 1995), supports it as the largest theropod ever found" (pdf). Gaia Revista de Geociências 15: 117–122. http://www.mnhn.ul.pt/geologia/gaia/7.pdf.
- ^ a b c d e f Brochu, C.R. (2003). "Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull". Society of Vertebrate Paleontology Memoirs 7: 1–138. doi:10.2307/3889334. JSTOR 3889334.
- ^ Lipkin, Christine; and Carpenter, Kenneth (2008). "Looking again at the forelimb of Tyrannosaurus rex". In Carpenter, Kenneth; and Larson, Peter E. (editors). Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. pp. 167–190. ISBN 0-253-35087-5.
- ^ "Museum unveils world's largest T-rex skull" (Press release). Montana State University. 2006-04-07. http://www.montana.edu/cpa/news/nwview.php?article=3607. Retrieved 2008-09-13.
- ^ a b Stevens, Kent A. (June 2006). "Binocular vision in theropod dinosaurs" (PDF). Journal of Vertebrate Paleontology 26 (2): 321–330. doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2. Archived from the original on 2008-02-16. http://web.archive.org/web/20080216074425/http://www.geocities.com/Athens/Bridge/4602/theropod_binocularvision.pdf.
- ^ a b Jaffe, Eric (2006-07-01). "Sight for 'Saur Eyes: T. rex vision was among nature's best". Science News 170 (1): 3. doi:10.2307/4017288. JSTOR 4017288. http://www.sciencenews.org/view/generic/id/7500/title/Sight_for_Saur_Eyes_%3Ci%3ET._rex%3Ci%3E_vision_was_among_natures_best. Retrieved 2008-10-06.
- ^ Snively, Eric; Donald M. Henderson, and Doug S. Phillips (2006). "Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for cranial strength and feeding mechanics" (PDF). Acta Palaeontologica Polonica 51 (3): 435–454. http://www.app.pan.pl/archive/published/app51/app51-435.pdf. Retrieved 2008-10-08.
- ^ Erickson, G.M.; Van Kirk, S.D.; Su, J.; Levenston, M.E.; Caler, W.E.; and Carter, D.R. (1996). "Bite-force estimation for Tyrannosaurus rex from tooth-marked bones". Nature 382 (6593): 706–708. doi:10.1038/382706a0.
- ^ Meers, M.B. (August 2003). "Maximum bite force and prey size of Tyrannosaurus rex and their relationships to the inference of feeding behavior". Historical Biology: A Journal of Paleobiology 16 (1): 1–12. doi:10.1080/0891296021000050755. http://www.ingentaconnect.com/content/tandf/ghbi/2003/00000016/00000001/art00001.
- ^ a b c Holtz, Thomas R. (1994). "The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics". Journal of Palaeontology 68 (5): 1100–1117. http://www.jstor.org/pss/1306180. Retrieved 2008-10-08.
- ^ a b c d Paul, Gregory S. (1988). Predatory dinosaurs of the world: a complete illustrated guide. New York: Simon and Schuster. ISBN 0-671-61946-2. OCLC 18350868.[page needed]
- ^ Smith, J.B. (December 2005). "Heterodonty in Tyrannosaurus rex: implications for the taxonomic and systematic utility of theropod dentitions" (PDF). Journal of Vertebrate Paleontology 25 (4): 865–887. doi:10.1671/0272-4634(2005)025[0865:HITRIF]2.0.CO;2. Archived from the original on 2008-02-16. http://web.archive.org/web/20080216074427/http://www.geocities.com/Athens/Bridge/4602/trexteeth.pdf.
- ^ Douglas K, Young S (1998). "The dinosaur detectives". New Scientist. http://www.newscientist.com/channel/life/dinosaurs/mg15821305.300. Retrieved 2008-10-16. "One palaeontologist memorably described the huge, curved teeth of T. rex as 'lethal bananas'"
- ^ a b Currie, Philip J.; Jørn H. Hurum and Karol Sabath (2003). "Skull structure and evolution in tyrannosaurid dinosaurs" (PDF). Acta Palaeontologica Polonica 48 (2): 227–234. http://www.app.pan.pl/archive/published/app48/app48-227.pdf. Retrieved 2008-10-08.
- ^ a b c Holtz, Thomas R., Jr. (2004). "Tyrannosauroidea". In David B. Weishampel, Peter Dodson and Halszka Osmólska. The dinosauria. Berkeley: University of California Press. pp. 111–136. ISBN 0-520-24209-2.
- ^ Maleev, E. A. (1955). "[Gigantic carnivorous dinosaurs of Mongolia]" (in Russian). Doklady Akademii Nauk SSSR 104 (4): 634–637.
- ^ Rozhdestvensky, AK (1965). "Growth changes in Asian dinosaurs and some problems of their taxonomy". Paleontological Journal 3: 95–109.
- ^ Carpenter, Kenneth (1992). "Tyrannosaurids (Dinosauria) of Asia and North America". In Niall J. Mateer and Pei-ji Chen. Aspects of nonmarine Cretaceous geology. Beijing: China Ocean Press. ISBN 9787502714635. OCLC 28260578.
- ^ Carr, Thomas D.; Thomas E. Williamson and David R. Schwimmer (March 2005). "A New Genus and Species of Tyrannosauroid from the Late Cretaceous (Middle Campanian) Demopolis Formation of Alabama". Journal of Vertebrate Paleontology 25 (1): 119–143. doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2.
- ^ Hurum, Jørn H.; Karol Sabath (2003). "Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared" (PDF). Acta Palaeontologica Polonica 48 (2): 161–190. http://www.app.pan.pl/archive/published/app48/app48-161.pdf. Retrieved 2008-10-08.
- ^ Olshevsky, George (1995). "The origin and evolution of the tyrannosaurids". Kyoryugaku Saizensen [Dino Frontline] 9–10: 92–119.
- ^ Carr, T.D.; T.E. Williamson (2004). "Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America". Zoological Journal of the Linnean Society 142: 479–523. doi:10.1111/j.1096-3642.2004.00130.x.
- ^ Gilmore, C.W. (1946). "A new carnivorous dinosaur from the Lance Formation of Montana". Smithsonian Miscellaneous Collections 106: 1–19.
- ^ Bakker, R.T.; M. Williams and P.J. Currie (1988). "Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana". Hunteria 1 (5): 1–30.
- ^ Carr, TD (1999). "Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Theropoda)". Journal of Vertebrate Paleontology 19: 497–520. doi:10.1080/02724634.1999.10011161.
- ^ Currie, Philip J. (2003). "Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada" (PDF). Acta Palaeontologica Polonica 42 (2): 191–226. http://www.app.pan.pl/archive/published/app48/app48-191.pdf. Retrieved 2008-10-09.
- ^ a b Breithaupt, Brent H.; Elizabeth H. Southwell and Neffra A. Matthews (2005-10-18). "In Celebration of 100 years of Tyrannosaurus Rex: Manospondylus Gigas, Ornithomimus Grandis, and Dynamosaurus Imperiosus, the Earliest Discoveries of Tyrannosaurus Rex in the West". Abstracts with Programs. 37. 2005 Salt Lake City Annual Meeting. Geological Society of America. pp. 406. http://gsa.confex.com/gsa/2005AM/finalprogram/abstract_96150.htm. Retrieved 2008-10-08.
- ^ a b Osborn, H. F. (1917). "Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus". Bulletin of the American Museum of Natural History (New York City: American Museum of Natural History) 35 (43): 733–771. http://hdl.handle.net/2246/1334. Retrieved 2008-10-08.
- ^ Anonymous, 2000. "Discovery could Endanger T.Rex Name." The Associated Press. June 13, 2000.
- ^ Ride, W. D. L. (1999). "Article 23.9 – Reversal of Precedence". International code of zoological nomenclature. London: International Commission on Zoological Nomenclature. ISBN 0-85301-006-4. OCLC 183090345. http://www.iczn.org/iczn/index.jsp?article=23&nfv=#9. Retrieved 2008-10-08.
- ^ Taylor, Mike (2002-08-27). "So why hasn't Tyrannosaurus been renamed Manospondylus?". The Dinosaur FAQ. http://www.miketaylor.org.uk/dino/faq/s-class/priority/. Retrieved 2008-10-08.
- ^ a b Horner JR, Padian K (September 2004). "Age and growth dynamics of Tyrannosaurus rex". Proceedings. Biological sciences / the Royal Society 271 (1551): 1875–80. doi:10.1098/rspb.2004.2829. PMC 1691809. PMID 15347508. http://journals.royalsociety.org/openurl.asp?genre=article&issn=0962-8452&volume=271&issue=1551&spage=1875. Retrieved 2008-10-05.
- ^ a b Schweitzer MH, Wittmeyer JL, Horner JR (June 2005). "Gender-specific reproductive tissue in ratites and Tyrannosaurus rex". Science 308 (5727): 1456–60. doi:10.1126/science.1112158. PMID 15933198. http://www.sciencemag.org/cgi/pmidlookup?view=long&pmid=15933198. Retrieved 2008-10-05.
- ^ Lee, Andrew H.; and Werning, Sarah (2008). "Sexual maturity in growing dinosaurs does not fit reptilian growth models". Proceedings of the National Academy of Sciences 105 (2): 582–587. doi:10.1073/pnas.0708903105. PMC 2206579. PMID 18195356. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2206579.
- ^ a b Erickson GM, Currie PJ, Inouye BD, Winn AA (July 2006). "Tyrannosaur life tables: an example of nonavian dinosaur population biology". Science 313 (5784): 213–7. doi:10.1126/science.1125721. PMID 16840697.
- ^ Carpenter, Kenneth (1992). "Variation in Tyrannosaurus rex". In Kenneth Carpenter and Philip J. Currie. Dinosaur Systematics: Approaches and Perspectives. Cambridge: Cambridge University Press. pp. 141–145. ISBN 0-521-43810-1.
- ^ Larson, P.L. 1994. Tyrannosaurus sex. In: Rosenberg, G.D. & Wolberg, D.L. Dino Fest. The Paleontological Society Special Publications. 7: 139–155.
- ^ Erickson GM, Kristopher Lappin A, Larson P (2005). "Androgynous rex – the utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs". Zoology (Jena, Germany) 108 (4): 277–86. doi:10.1016/j.zool.2005.08.001. PMID 16351976. http://linkinghub.elsevier.com/retrieve/pii/S0944-2006(05)00047-4. Retrieved 2008-10-08.
- ^ Schweitzer MH, Elsey RM, Dacke CG, Horner JR, Lamm ET (April 2007). "Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone?". Bone 40 (4): 1152–8. doi:10.1016/j.bone.2006.10.029. PMID 17223615. http://linkinghub.elsevier.com/retrieve/pii/S8756-3282(06)00748-4. Retrieved 2008-10-08.
- ^ Leidy, J (1865). "Memoir on the extinct reptiles of the Cretaceous formations of the United States". Smithsonian Contributions to Knowledge 14: 1–135.
- ^ "Tyrannosaurus". American Museum of Natural History. http://www.amnh.org/exhibitions/expeditions/treasure_fossil/Treasures/Tyrannosaurus/tyrannos.html?dinos. Retrieved 2008-10-16.
- ^ a b Newman, BH (1970). "Stance and gait in the flesh-eating Tyrannosaurus". Biological Journal of the Linnean Society 2: 119–123. doi:10.1111/j.1095-8312.1970.tb01707.x.
- ^ "The Age of Reptiles Mural". Yale University. 2008. http://www.yale.edu/peabody/explore/reptiles.html. Retrieved 2008-10-16.
- ^ a b c Osborn, H. F. (1905). "Tyrannosaurus and other Cretaceous carnivorous dinosaurs". Bulletin of the AMNH (New York City: American Museum of Natural History) 21 (14): 259–265. http://hdl.handle.net/2246/1464. Retrieved 2008-10-06.
- ^ Lambe, L. M. (1914). "On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon". Ottawa Naturalist 27: 129–135.
- ^ a b c d Horner, John R.; Don Lessem (1993). The complete T. rex. New York City: Simon & Schuster. ISBN 0-671-74185-3.[page needed]
- ^ a b Osborn, Henry Fairfield; Barnum Brown (1906). "Tyrannosaurus, Upper Cretaceous carnivorous dinosaur". Bulletin of the AMNH (New York City: American Museum of Natural History) 22 (16): 281–296. http://hdl.handle.net/2246/1473. Retrieved 2008-10-06.
- ^ Carpenter, Kenneth; Matt Smith (2001). "Forelimb Osteology and Biomechanics of Tyrannosaurus rex". In Darren Tanke and Kenneth Carpenter. Mesozoic vertebrate life. Bloomington: Indiana University Press. pp. 90–116. ISBN 0-253-33907-3.
- ^ In http://users.tamuk.edu/kfjab02/dinos/VPTHEROPOD.htm , text search for ARCTOMETATARSALIA".
- ^ a b Fields, Helen (May 2006). "Dinosaur Shocker". Smithsonian Magazine. http://www.smithsonianmag.com/science-nature/10021606.html. Retrieved 2008-10-02.
- ^ Schweitzer, Mary H.; Jennifer L. Wittmeyer, John R. Horner and Jan K. Toporski (March 2005). "Soft-tissue vessels and cellular preservation in Tyrannosaurus rex". Science 307 (5717): 1952–5. Bibcode 2005Sci...307.1952S. doi:10.1126/science.1108397. PMID 15790853. http://www.sciencemag.org/cgi/content/abstract/307/5717/1952. Retrieved 2008-10-02.
- ^ Rincon, Paul (2007-04-12). "Protein links T. rex to chickens". BBC News. http://news.bbc.co.uk/2/hi/science/nature/6548719.stm. Retrieved 2008-10-02.
- ^ Vergano, Dan (2007-04-13). "Yesterday's T. Rex is today's chicken". USA Today. http://www.usatoday.com/tech/science/discoveries/2007-04-12-trex-protein_N.htm. Retrieved 2008-10-08.
- ^ Schmid, Randolph E.; Associated Press (2008-04-24). "Scientists study evidence modern birds came from dinosaurs". Newsvine. http://www.newsvine.com/_news/2008/04/24/1450460-scientists-study-evidence-modern-birds-came-from-dinosaurs. Retrieved 2008-10-08.
- ^ Kaye, Thomas G.; Gary Gaugler and Zbigniew Sawlowicz (July 2008). Stepanova, Anna. ed. "Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms". ONE 3 (7): e2808. doi:10.1371/journal.pone.0002808. PMC 2483347. PMID 18665236. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2483347. Retrieved 2008-07-29.
- ^ "New Research Challenges Notion That Dinosaur Soft Tissues Still Survive" (Press release). Newswise. 2008-07-24. http://newswise.com/articles/view/542898/. Retrieved 2008-10-08.
- ^ "Researchers Debate: Is It Preserved Dinosaur Tissue, or Bacterial Slime?" (Press release). Discover. 2008-07-30. http://blogs.discovermagazine.com/80beats/2008/07/30/researchers-debate-is-it-preserved-dinosaur-tissue-or-bacterial-slime/. Retrieved 2008-09-04.
- ^ San Antonio JD, Schweitzer MH, Jensen ST, Kalluri R, Buckley M, et al. (2011). "Dinosaur peptides suggest mechanisms of protein survival". PLoS ONE 6: e20381. doi:10.1371/journal.pone.0020381. http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0020381.
- ^ Paul, Gregory S. (2008). "The extreme lifestyles and habits of the gigantic tyrannosaurid superpredators of the Late Cretaceous of North America and Asia". In Carpenter, Kenneth; and Larson, Peter E. (editors). Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. p. 316. ISBN 0-253-35087-5.
- ^ Xu, Xing; Mark A. Norell, Xuewen Kuang, Xiaolin Wang, Qi Zhao and Chengkai Jia (2004-10-07). "Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids". Nature 431 (7009): 680–684. doi:10.1038/nature02855. PMID 15470426.
- ^ Bakker, Robert T. (1968). "The superiority of dinosaurs" (PDF). Discovery 3 (2): 11–12. Archived from the original on September 9, 2006. http://web.archive.org/web/20060909131058/http://bio.fsu.edu/~amarquez/Evolutionary%20Morphology%20fall%202004/Bakker/Bakker%201968%20-%20Superiority%20of%20DInos.pdf. Retrieved 2008-10-07.
- ^ Bakker, Robert T. (1972). "Anatomical and ecological evidence of endothermy in dinosaurs" (PDF). Nature 238 (5359): 81–85. Bibcode 1972Natur.238...81B. doi:10.1038/238081a0. Archived from the original on September 9, 2006. http://web.archive.org/web/20060909173036/http://bio.fsu.edu/~amarquez/Evolutionary%20Morphology%20fall%202004/Bakker/14-%20Bakker%201972%20-%20dino%20endothermy.pdf. Retrieved 2008-10-07.
- ^ Barrick, Reese E.; William J. Showers (July 1994). "Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes". Science (New York City) 265 (5169): 222–224. doi:10.1126/science.265.5169.222. PMID 17750663. http://www.sciencemag.org/cgi/content/abstract/265/5169/222. Retrieved 2008-10-07.
- ^ Trueman, Clive; Carolyn Chenery, David A. Eberth and Baruch Spiro (2003). "Diagenetic effects on the oxygen isotope composition of bones of dinosaurs and other vertebrates recovered from terrestrial and marine sediments". Journal of the Geological Society 160 (6): 895. doi:10.1144/0016-764903-019.
- ^ Barrick, Reese E.; William J. Showers (October 1999). "Thermophysiology and biology of Giganotosaurus: comparison with Tyrannosaurus". Palaeontologia Electronica 2 (2). http://palaeo-electronica.org/1999_2/gigan/issue2_99.htm. Retrieved 2008-10-07.
- ^ Barrick, Reese E.; Michael K. Stoskopf and William J. Showers (1999). "Oxygen isotopes in dinosaur bones". In James O. Farlow and M. K. Brett-Surman. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 474–490. ISBN 0-253-21313-4.
- ^ Paladino, Frank V.; James R. Spotila and Peter Dodson (1999). "A blueprint for giants: modeling the physiology of large dinosaurs". In James O. Farlow and M. K. Brett-Surman. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 491–504. ISBN 0-253-21313-4.
- ^ Chinsamy, Anusuya; Willem J. Hillenius (2004). "Physiology of nonavian dinosaurs". In David B. Weishampel, Peter Dodson and Halszka Osmólska. The dinosauria. Berkeley: University of California Press. pp. 643–659. ISBN 0-520-24209-2.
- ^ Lockley, MG; Hunt AP (1994). "A track of the giant theropod dinosaur Tyrannosaurus from close to the Cretaceous/Tertiary boundary, northern New Mexico". Ichnos 3: 213–218. doi:10.1080/10420949409386390.
- ^ "T.rex footprint discovered". Natural History Museum. 2007. http://www.nhm.ac.uk/about-us/news/2007/october/news_12515.html. Retrieved 2008-12-09.
- ^ Hutchinson JR, Ng-Thow-Hing V, Anderson FC (June 2007). "A 3D interactive method for estimating body segmental parameters in animals: application to the turning and running performance of Tyrannosaurus rex". Journal of theoretical biology 246 (4): 660–80. doi:10.1016/j.jtbi.2007.01.023. PMID 17363001.
- ^ Carrier, David R.; Rebecca M. Walter and David V. Lee (2001-11-15). "Influence of rotational inertia on turning performance of theropod dinosaurs: clues from humans with increased rotational inertia". Journal of Experimental Biology (Company of Biologists) 204 (22): 3917–3926. PMID 11807109. http://jeb.biologists.org/cgi/content/full/204/22/3917.
- ^ a b Hutchinson, J.R. (2004). "Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa" (PDF). Journal of Morphology 262 (1): 441–461. doi:10.1002/jmor.10240. PMID 15352202. http://www.rvc.ac.uk/AboutUs/Staff/jhutchinson/documents/JRH13.pdf.
- ^ a b c d e Hutchinson JR, Garcia M (February 2002). "Tyrannosaurus was not a fast runner". Nature 415 (6875): 1018–21. doi:10.1038/4151018a. PMID 11875567.
- ^ Holtz, Thomas R. (1996-05-01). "Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda)". Journal of Paleontology 70 (3): 536–538. http://jpaleontol.geoscienceworld.org/cgi/content/abstract/70/3/536. Retrieved 2008-10-03.
- ^ Christiansen, P. (1998). "Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential" (pdf). Gaia 15: 241–255. ISSN 0871-5424. http://www.mnhn.ul.pt/geologia/gaia/19.pdf.
- ^ "Giraffe". WildlifeSafari.info. http://www.wildlifesafari.info/giraffe.html. Retrieved 2006-04-29.
- ^ "The History of Woodland Park Zoo – Chapter 4". http://www.zoo.org/zoo_info/history/chapters/history4.html. Retrieved 2006-04-29.
- ^ Alexander, R.M. (August 7, 2006). "Dinosaur biomechanics". Proc Biol Sci. (The Royal Society) 273 (1596): 1849–1855. doi:10.1098/rspb.2006.3532. PMC 1634776. PMID 16822743. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1634776.
- ^ a b Hanna, Rebecca R. (2002). "Multiple injury and infection in a sub-adult theropod dinosaur (Allosaurus fragilis) with comparisons to allosaur pathology in the Cleveland-Lloyd dinosaur quarry collection". Journal of Vertebrate Paleontology 22 (1): 76–90. doi:10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2. catalogs the injuries of the Allosaurus known as "Big Al" – at least one was attributed to a fall.
- ^ Sellers, W.I., and Manning, P.L. (July 2007). "Estimating dinosaur maximum running speeds using evolutionary robotics". Proc. R. Soc. B (The Royal Society) 274 (1626): 2711. doi:10.1098/rspb.2007.0846. PMC 2279215. PMID 17711833. http://dinonews.net/rubriq/articles.php5?action=open&ref=2007_sellers_speed.
- ^ Seward, L (2007-08-21). "T. rex 'would outrun footballer'". BBCNews. http://news.bbc.co.uk/2/hi/science/nature/6956867.stm. Retrieved 2008-10-16.
- ^ Callison, G.; H. M. Quimby (1984). "Tiny dinosaurs: Are they fully grown?". Journal of Vertebrate Paleontology 3: 200–209. doi:10.1080/02724634.1984.10011975.
- ^ a b Manning P (2008). "T. rex speed trap". In Carpenter, Kenneth; Larson, Peter E.. Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. pp. 205–228. ISBN 0-253-35087-5.
- ^ Paul, G.S., and Christiansen, P. (September 2000). "Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion". Paleobiology 26 (3): 450. doi:10.1666/0094-8373(2000)026<0450:FPINDI>2.0.CO;2. http://www.bioone.org/perlserv/?request=get-document&doi=10.1666%2F0094-8373(2000)026%3C0450%3AFPINDI%3E2.0.CO%3B2.
- ^ Lambe, L. B. (1917). "The Cretaceous theropodous dinosaur Gorgosaurus". Memoirs of the Geological Survey of Canada 100: 1–84.
- ^ Farlow, J. O. and Holtz, T. R. (2002). "The Fossil Record of Predation". In Kowalewski, M. and Kelley, P.H. (pdf). pp. 251–266. http://www.yale.edu/ypmip/predation/Chapter_09.pdf.
- ^ a b c Horner, J.R. (1994). "Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger)". The Paleontological Society Special Publication 7: 157–164.
- ^ Amos, J. (2003-07-31). "T. rex goes on trial". BBC. http://news.bbc.co.uk/1/hi/sci/tech/3112527.stm.
- ^ Novella, S. "Interview with Jack Horner." The Skeptics Guide to the Universe. 14-OCT-2011. Accessed 24-OCT-2011, http://media.libsyn.com/media/skepticsguide/skepticast2009-10-14.mp3
- ^ The king of killers or fearsome freeloader? It is one of the big questions currently in palaeontology.
- ^ "T. Rex brain study reveals a refined "nose"". Calgary Herald. 2008-10-28. http://www.canada.com/calgaryherald/story.html?id=3641f27e-ca2e-44e8-a56b-f9a0b4aef4b5. Retrieved 2008-10-29.
- ^ Paul, G.S. (1988). Predatory Dinosaurs of the World. Simon and Schuster. ISBN 0-671-61946-2. OCLC 18350868.
- ^ Ruxton GD, Houston DC (April 2003). "Could Tyrannosaurus rex have been a scavenger rather than a predator? An energetics approach". Proceedings. Biological sciences / the Royal Society 270 (1516): 731–3. doi:10.1098/rspb.2002.2279. PMC 1691292. PMID 12713747. http://journals.royalsociety.org/openurl.asp?genre=article&issn=0962-8452&volume=270&issue=1516&spage=731. Retrieved 2008-10-05.
- ^ Chin, K., Erickson, G.M. et al. (1998-06-18). "A king-sized theropod coprolite". Nature 393 (6686): 680. doi:10.1038/31461. http://www.nature.com/nature/journal/v393/n6686/abs/393680a0.html. Summary at Monastersky, R. (1998-06-20). "Getting the scoop from the poop of T. rex". Science News 153 (25): 391. doi:10.2307/4010364. JSTOR 4010364. http://www.sciencenews.org/pages/sn_arc98/6_20_98/fob2.htm.
- ^ a b Walters, Martin (1995). Bloomsbury Illustrated Dictionary of Prehistoric Life (Bloomsbury Illustrated Dictionaries). Godfrey Cave Associates Ltd. ISBN 1-85471-648-4.
- ^ a b Wolff EDS, Salisbury SW, Horner JR, Varricchi DJ. "Common Avian Infection Plagued the Tyrant Dinosaurs". p. e7288. doi:10.1371/journal.pone.0007288. http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0007288;jsessionid=46F0A1EFB5E7F840BE2194BA7F0AEC02. Retrieved 2009-11-01.
- ^ Carpenter, K. (1998). "Evidence of predatory behavior by theropod dinosaurs". Gaia 15: 135–144. http://vertpaleo.org/publications/jvp/15-576-591.cfm. Retrieved 2007-12-05.
- ^ Happ, John; and Carpenter, Kenneth (2008). "An analysis of predator–prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops". In Carpenter, Kenneth; and Larson, Peter E. (editors). Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. pp. 355–368. ISBN 0-253-35087-5.
- ^ Tanke, Darren H.; and Currie, Philip J. (1998). "Head-biting behavior in theropod dinosaurs: paleopathological evidence" (pdf). Gaia (15): 167–184. ISSN 0871-5424. http://www.mnhn.ul.pt/geologia/gaia/12.pdf.
- ^ Carbone, Chris; Turvey, Samuel T.; Bielby, Jon (January 26, 2011). "Intra-guild Competition and its Implications for One of the Biggest Terrestrial Predators, Tyrannosaurus rex". Proceedings of the Royal Society B: Biological Sciences. doi:10.1098/rspb.2010.2497.
- ^ 1999. The teeth of the Tyrannosaurus. Scientific American 281: 40-41.
- ^ The Complete T. Rex: How Stunning New Discoveries are Changing our Understanding of the World's Most Famous Dinosaur copyright 1993 by John R. Horner: pp 214-215
- ^ a b Longrich N R., Horner J.R., Erickson G.M. & Currie P.J. (2010), "Cannibalism in Tyrannosaurus rex", Public Library of Science.
- ^ Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 331-336.
- ^ "The First Tyrannosaurus Skeleton, 1905". Linda Hall Library of Science, Engineering and Technology. Archived from the original on September 28, 2006. http://web.archive.org/web/20060928114635/http://www.lindahall.org/events_exhib/exhibit/exhibits/dino/osb1905.htm. Retrieved 2008-08-03.
- ^ Compare Oedipus rex, translated as Oedipus the King or King Oedipus, as the second part of the species name is syntactically an apposition to the genus name.
- ^ Breithaup, BH; Southwell EH, Matthews NA (2006). "Dynamosaurus imperiosus and the earliest discoveries of Tyrannosaurus rex in Wyoming and the West". New Mexico Museum of Natural History and Science Bulletin 35: 257–258.
- ^ "Footprint of a Giant". Online guide to the continental Cretaceous–Tertiary boundary in the Raton basin, Colorado and New Mexico. United States Geological Survey. http://esp.cr.usgs.gov/info/kt/footprint.html. Retrieved 2008-10-09.
- ^ "Preparation and mounting". Sue at the Field Museum. The Field Museum. 2007. http://www.fieldmuseum.org/sue/about_prep.asp. Retrieved 11 February 2010.
- ^ Erickson, G., Makovicky, P. J., Currie, P. J., Norell, M., Yerby, S., Brochu, C. A. (26 May 2004). "Gigantism and life history parameters of tyrannosaurid dinosaurs". Nature 430 (7001): 772–775. doi:10.1038/nature02699. PMID 15306807. http://www.nature.com/nature/journal/v430/n7001/abs/nature02699.html.
- ^ Brochu, C.A. (December 2003). "Lessons From A Tyrannosaur: The Ambassadorial Role Of Paleontology". PALAIOS 18 (6): 475. doi:10.1669/0883-1351(2003)018<0475:LFATTA>2.0.CO;2. http://www.bioone.org/perlserv/?request=get-document&doi=10.1669%2F0883-1351(2003)018%3C0475%3ALFATTA%3E2.0.CO%3B2.
- ^ [1]
- ^ Larson, Neal L. (2008). "One hundred years of Tyrannosaurus rex: the skeletons". In Larson, Peter; and Carpenter, Kenneth, editors. Tyrannosaurus rex, the tyrant king. Bloomington, IN: Indiana University Press. pp. 1–55. ISBN 978-0-253-35087-9.
- ^ Steve Fiffer (2000). Tyrannosaurus Sue. W. H. Freeman and Company, New York. ISBN 0-7167-4017-6. chapter 7 "Jurassic Farce", pp 121–2
- ^ "Dig pulls up five T. rex specimens". BBC News. 10 October 2000. http://news.bbc.co.uk/2/hi/science/nature/965609.stm. Retrieved 13 December 2008.
- ^ Currie, PJ; Huru, JH, Sabath, K (2003). "Skull structure and evolution in tyrannosaurid dinosaurs" (PDF). Acta Palaeontologica Polonica 48 (2): 227–234. http://www.biology.ualberta.ca/faculty/philip_currie/uploads/pdfs/2003/2003TyrannosauridEvolution.pdf. Retrieved 2008-10-16.
- ^ Henderson, M (In press). "Nano No More: The death of the pygmy tyrant". In Henderson, M. The origin, systematics, and paleobiology of Tyrannosauridae. Dekalb, Illinois: Northern Illinois University Press.
- ^ "Visit Jane the Dinosaur at the Burpee Museum, Rockford, Illinois". Archived from the original on May 25, 2008. http://web.archive.org/web/20080525153508/http://www.visitjane.com/index.cfm. Retrieved 2008-10-16.
- ^ "Museum unveils world's largest T-rex skull.". http://www.montana.edu/cpa/news/nwview.php?article=3607. Retrieved 2006-04-07.
- ^ Ryan, M. J. "New Biggest T-rex Skull.". http://palaeoblog.blogspot.com/2006/04/new-biggest-t-rex-skull.html. Retrieved 2006-04-12.
Further reading
- Farlow, J. O. (2000). "Theropod Locomotion". American Zoologist (The Society for Integrative and Comparative Biology) 40 (4): 640–663. doi:10.1093/icb/40.4.640. http://icb.oxfordjournals.org/cgi/content/full/40/4/640.
External links
- Tyrannosauridae "Tree of Life" page, very comprehensive survey by major authority Tom Holtz.
- Radford, Tim (12 August 2004). "The secret of T. rex's colossal size: a teenage growth spurt". London: The Guardian. http://www.guardian.co.uk/uk_news/story/0,3604,1281338,00.html. Retrieved 2010-04-09.
- Tyrannosaurus in the Dino Directory
- Sue's homepage
- Stan's homepage
- Unearthing Tyrannosaurus rex
- T.rex juvenile Jane
- NBCI's Taxonomy Browser
- Bristol University study on bite forces of predators
- Museum of Unnatural Mystery – Bite force etc. of T. rex
- Stanford University on bite force of T. rex
- How Tyrannosaurus might have had sex
- Recent Discovery of Soft Tissue
- Cretaceous Hell Creek Faunal Facies is an example of one tyrannosaur environment, in the Hell Creek Formation of Montana
- Miniature Tyrannosaurus Rex discovered
Categories:- Cretaceous dinosaurs
- Dinosaurs of North America
- Tyrannosaurs
- Geology of Montana
- Manospondylus

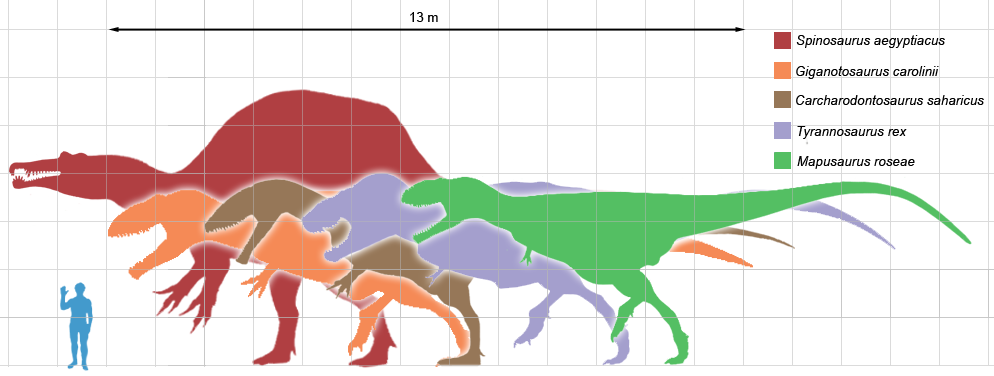






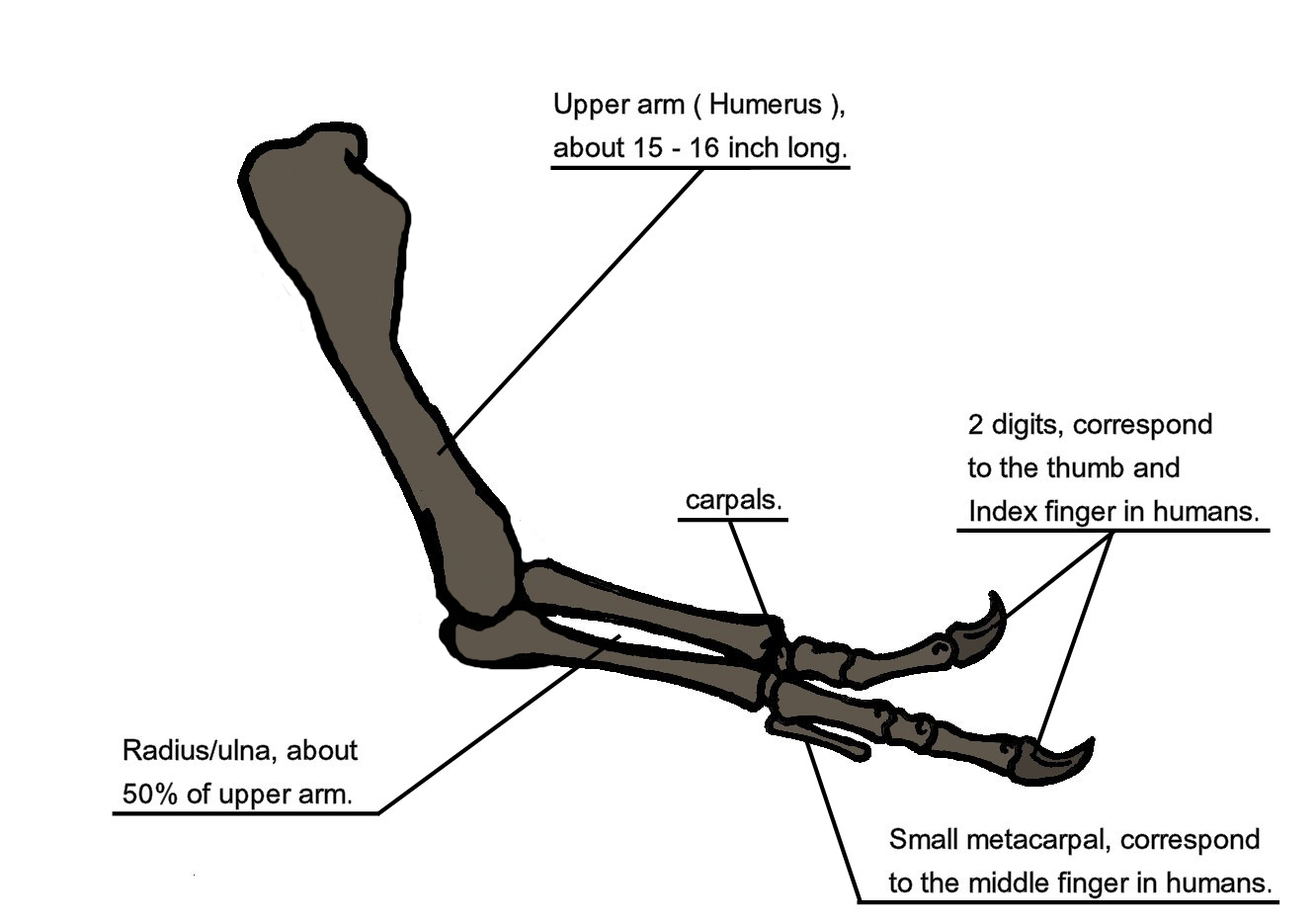





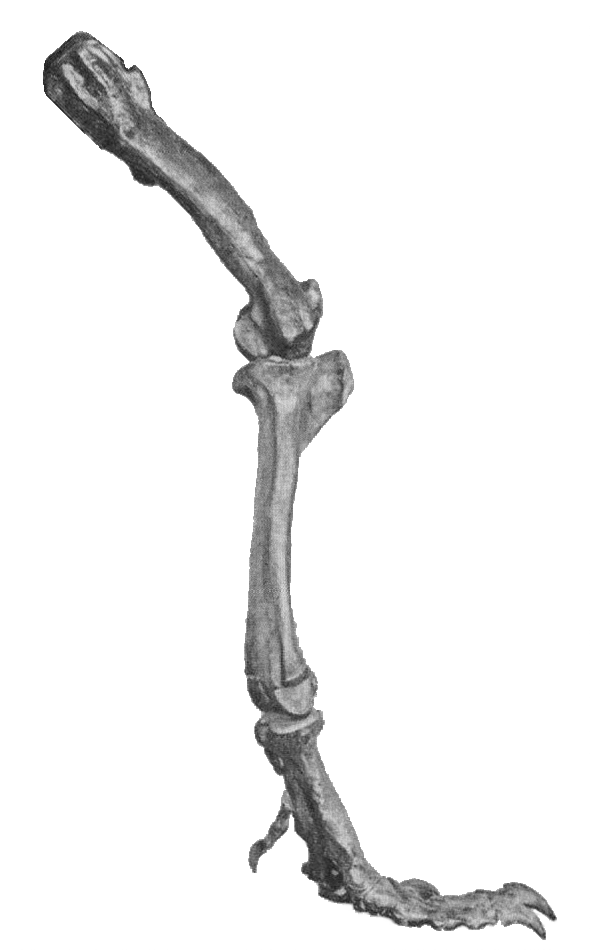



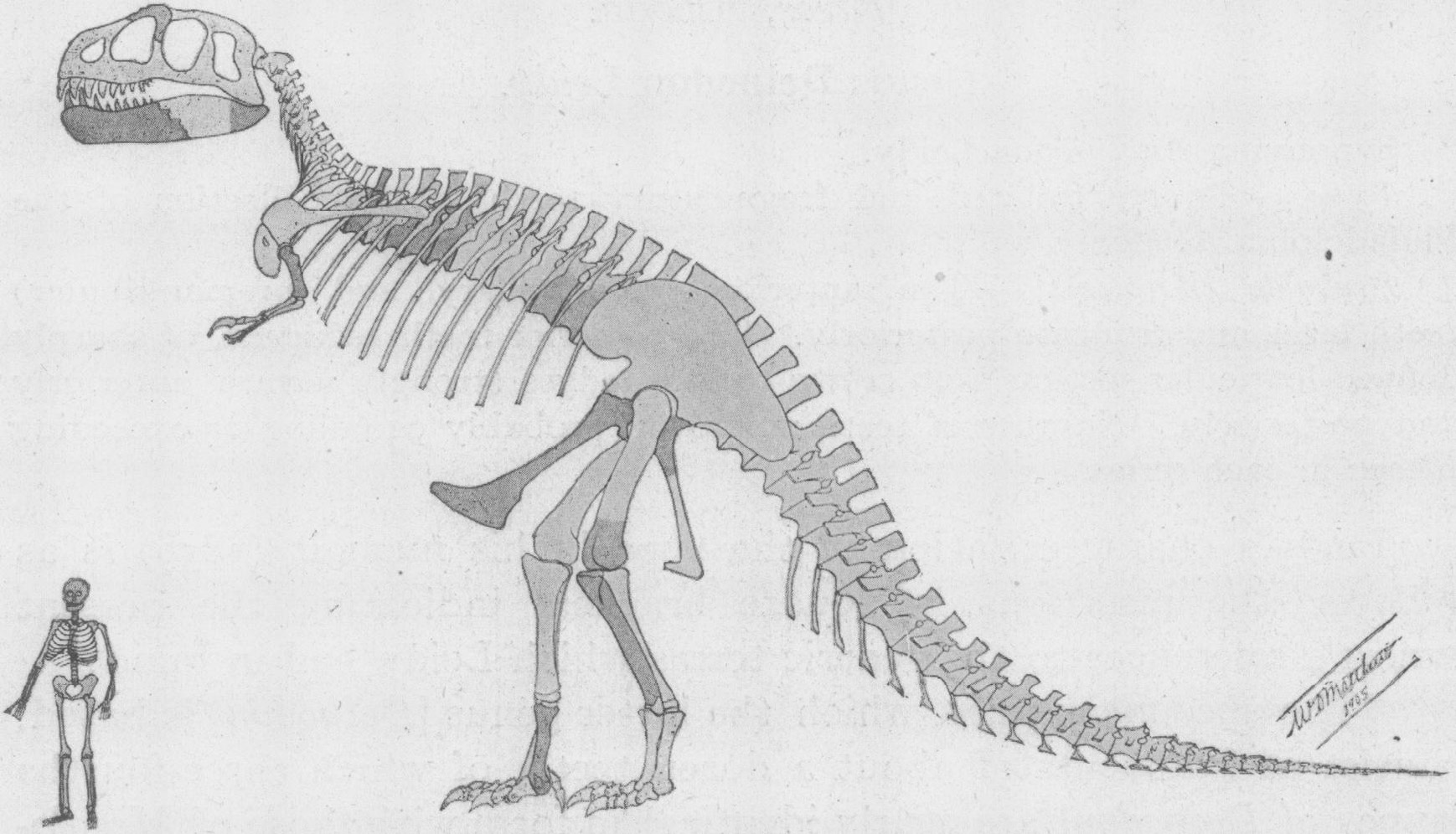


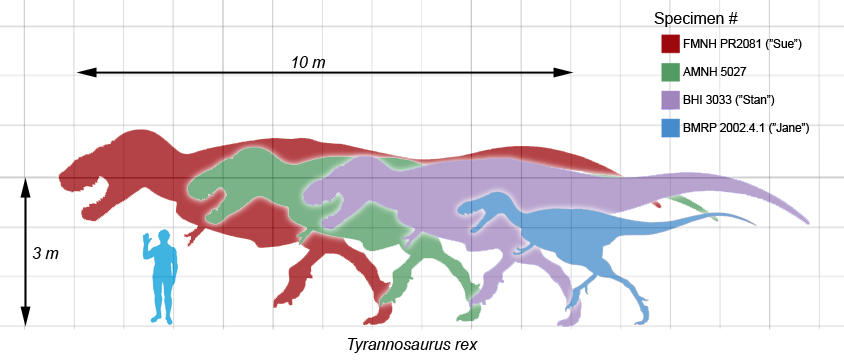
Wikimedia Foundation. 2010.