- NMDA receptor
-
 Stylised depiction of an activated NMDAR. Glutamate is in the glutamate binding site and glycine is in the glycine binding site. Allosteric sites that would cause inhibition of the receptor are not occupied. NMDARs require the binding of two molecules of glutamate or aspartate and two of glycine.[1]
Stylised depiction of an activated NMDAR. Glutamate is in the glutamate binding site and glycine is in the glycine binding site. Allosteric sites that would cause inhibition of the receptor are not occupied. NMDARs require the binding of two molecules of glutamate or aspartate and two of glycine.[1]
The NMDA receptor (NMDAR), a glutamate receptor, is the predominant molecular device for controlling synaptic plasticity and memory function.[2]
The NMDAR is a specific type of ionotropic glutamate receptor. NMDA (N-methyl D-aspartate) is the name of a selective agonist that binds to NMDA receptors but not to other glutamate receptors. Activation of NMDA receptors results in the opening of an ion channel that is nonselective to cations with an equilibrium potential near 0 mV. A unique property of the NMDA receptor is its voltage-dependent activation, a result of ion channel block by extracellular Mg2+ ions. This allows the flow of Na+ and small amounts of Ca2+ ions into the cell and K+ out of the cell to be voltage-dependent.[3][4][5][6]
Calcium flux through NMDARs is thought to play a critical role in synaptic plasticity, a cellular mechanism for learning and memory. The NMDA receptor is distinct in two ways: First, it is both ligand-gated and voltage-dependent; second, it requires co-activation by two ligands - glutamate and glycine[7].
Contents
Structure
The NMDA receptor forms a heterotetramer between two NR1 and two NR2 subunits (the subunits are also called glutamate-binding NMDA receptor subunits or GluN for short); two obligatory NR1 subunits and two regionally localized NR2 subunits. A related gene family of NR3 A and B subunits have an inhibitory effect on receptor activity. Multiple receptor isoforms with distinct brain distributions and functional properties arise by selective splicing of the NR1 transcripts and differential expression of the NR2 subunits.
Each receptor subunit has modular design and each structural module also represents a functional unit:
- The extracellular domain contains two globular structures: a modulatory domain and a ligand-binding domain. NR1 subunits bind the co-agonist glycine and NR2 subunits bind the neurotransmitter glutamate.
- The agonist-binding module links to a membrane domain, which consists of three trans-membrane segments and a re-entrant loop reminiscent of the selectivity filter of potassium channels.
- The membrane domain contributes residues to the channel pore and is responsible for the receptor's high-unitary conductance, high-calcium permeability, and voltage-dependent magnesium block.
- Each subunit has an extensive cytoplasmic domain, which contain residues that can be directly modified by a series of protein kinases and protein phosphatases, as well as residues that interact with a large number of structural, adaptor, and scaffolding proteins.
The glycine-binding modules of the NR1 and NR3 subunits and the glutamate-binding module of the NR2A subunit have been expressed as soluble proteins, and their three-dimensional structure has been solved at atomic resolution by x-ray crystallography. This has revealed a common fold with amino acid-binding bacterial proteins and with the glutamate-binding module of AMPA-receptors and kainate-receptors.
Variants
NR1
There are eight variants of the NR1 subunit produced by alternative splicing of GRIN1:[8]
- NR1-1a, NR1-1b; NR1-1a is the most abundantly expressed form.
- NR1-2a, NR1-2b;
- NR1-3a, NR1-3b;
- NR1-4a, NR1-4b;
NR2
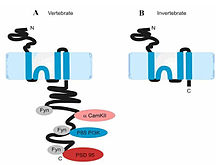 NR2 subunit in vertebrates (left) and invertebrates (right). Ryan et al., 2008
NR2 subunit in vertebrates (left) and invertebrates (right). Ryan et al., 2008While a single NR2 subunit is found in invertebrate organisms, four distinct isoforms of the NR2 subunit are expressed in vertebrates and are referred to with the nomenclature NR2A through D(coded byGRIN2A, GRIN2B, GRIN2C, GRIN2D). Strong evidence shows that the genes coding the NR2 subunits in vertebrates have undergone at least two rounds of gene duplication[9]. They contain the binding-site for the neurotransmitter glutamate. More importantly, each NR2 subunit has a different intracellular C-terminal domain that can interact with different sets signaling messengers[10]. Unlike NR1 subunits, NR2 subunits are expressed differentially across various cell types and control the electrophysiological properties of the NMDA receptor. One particular subunit, NR2B, is mainly present in immature neurons and in extrasynaptic locations, and contains the binding-site for the selective inhibitor ifenprodil.
Whereas NR2B is predominant in the early postnatal brain, the number of NR2A subunits grows, and eventually NR2A subunits outnumber NR2B. This is called NR2B-NR2A developmental switch, and is notable because of the different kinetics each NR2 subunit lends to the receptor.[11]
There are three hypothetical models to describe this switch mechanism:
- Dramatic increase in synaptic NR2A along with decrease in NR2B
- Extrasynaptic displacement of NR2B away from the synapse with increase in NR2A
- Increase of NR2A diluting the number of NR2B without the decrease of the latter.
The NR2B and NR2A subunits also have differential roles in mediating excitotoxic neuronal death.[12] The developmental switch in subunit composition is thought to explain the developmental changes in NMDA neurotoxicity.[13] Disruption of the gene for NR2B in mice causes perinatal lethality, whereas the disruption of NR2A gene produces viable mice, although with impaired hippocampal plasticity[14]. One study suggests that reelin may play a role in the NMDA receptor maturation by increasing the NR2B subunit mobility.[15]
NR2B to NR2C switch
Granule cell precursors (GCPs) of the cerebellum, after undergoing symmetric cell division[16] in the external granule-cell layer (EGL), migrate into the internal granule-cell layer (IGL) where they downregulate NR2B and activate NR2C, a process that is independent of neuregulin beta signaling through ErbB2 and ErbB4 receptors.[17]
Ligands
Agonists
Activation of NMDA receptors requires binding of glutamate or aspartate (aspartate does not stimulate the receptors as strongly).[18] In addition, NMDARs also require the binding of the co-agonist glycine for the efficient opening of the ion channel, which is a part of this receptor.
D-serine has also been found to co-agonize the NMDA receptor with even greater potency than glycine.[19] D-serine is produced by serine racemase, and is enriched in the same areas as NMDA receptors. Removal of D-serine can block NMDA-mediated excitatory neurotransmission in many areas. Recently, it has been shown that D-serine can be released both by neurons and astrocytes to regulate NMDA receptors.
In addition, a third requirement is membrane depolarization. A positive change in transmembrane potential will make it more likely that the ion channel in the NMDA receptor will open by expelling the Mg2+ ion that blocks the channel from the outside. This property is fundamental to the role of the NMDA receptor in memory and learning, and it has been suggested that this channel is a biochemical substrate of Hebbian learning, where it can act as a coincidence detector for membrane depolarization and synaptic transmission.
Known NMDA receptor agonists include:
- Aminocyclopropanecarboxylic acid
- D-Cycloserine
- cis-2,3-Piperidinedicarboxylic acid
- L- aspartate
- Quinolinate
- Homocysterate
- D-serine
- ACPL
- L-alanine
Antagonists
Main article: NMDA receptor antagonistAntagonists of the NMDA receptor are used as anesthetics for animals and sometimes humans, and are often used as recreational drugs due to their hallucinogenic properties, in addition to their unique effects at elevated dosages such as dissociation. When NMDA receptor antagonists are given to rodents in large doses, they can cause a form of brain damage called Olney's Lesions. So far, the published research on Olney's Lesions is inconclusive in its occurrence upon human or monkey brain tissues with respect to an increase in the presence of NMDA receptor antagonists.[20]
Common NMDA receptor antagonists include:
- Amantadine[21]
- Ketamine
- Phencyclidine (PCP)
- Nitrous oxide
- Dextromethorphan and dextrorphan
- Memantine
- Ethanol
- Riluzole (used in ALS)[22]
- Xenon
- HU-211 (also a cannabinoid)
- Lead (Pb2+)[23]
- Conantokins
Dual opioids and NMDA-Antagonists:
Modulators
The NMDA receptor is modulated by a number of endogenous and exogenous compounds:[24]
- Mg2+ not only blocks the NMDA channel in a voltage-dependent manner but also potentiates NMDA-induced responses at positive membrane potentials. Magnesium glycinate and magnesium taurinate treatment has been used to produce rapid recovery from depression.[25]
- Na+, K+ and Ca2+ not only pass through the NMDA receptor channel but also modulate the activity of NMDA receptors.
- Zn2+ and Cu2+ generally block NMDA current activity in a noncompetitive and a voltage-independent manner. However zinc may potentiate or inhibit the current depending on the neural activity. (Zinc and Copper Influence Excitability of Rat Olfactory Bulb Neurons by Multiple Mechanisms|http://jn.physiology.org/content/86/4/1652.short)
- Pb2+ lead is a potent NMDAR antagonist. Presynaptic deficits resulting from Pb2+ exposure during synaptogenesis are mediated by disruption of NMDAR-dependent BDNF signaling.
- It has been demonstrated that polyamines do not directly activate NMDA receptors, but instead act to potentiate or inhibit glutamate-mediated responses.
- Aminoglycosides have been shown to have a similar effect to polyamines, and this may explain their neurotoxic effect.
- The activity of NMDA receptors is also strikingly sensitive to the changes in H+ concentration, and partially inhibited by the ambient concentration of H+ under physiological conditions.[citation needed] The level of inhibition by H+ is greatly reduced in receptors containing the NR1a subtype, which contains the positively-charged insert Exon 5. The effect of this insert may be mimicked by positively-charged polyamines and aminoglycosides, explaining their mode of action.
- NMDA receptor function is also strongly regulated by chemical reduction and oxidation, via the so-called "redox modulatory site."[26] Through this site, reductants dramatically enhance NMDA channel activity, whereas oxidants either reverse the effects of reductants or depress native responses. It is generally believed that NMDA receptors are modulated by endogenous redox agents such as glutathione, lipoic acid, and the essential nutrient pyrroloquinoline quinone.[27]
- Src kinase enhances NMDA receptor currents.[28]
- Reelin modulates NMDA function through Src family kinases and DAB1.[29] significantly enhancing LTP in the hippocampus.
- CDK5 regulates the amount of NR2B-containing NMDA receptors on the synaptic membrane, thus affecting synaptic plasticity.[30][31]
Functional role
The NMDA receptor is a non-specific cation channel which can allow Ca2+, Na+, and K+ to pass into the cell. The excitatory postsynaptic potential (EPSP) produced by activation of an NMDA receptor increases the concentration of Ca2+ in the cell. The Ca2+ can in turn function as a second messenger in various signaling pathways. However, the NMDA receptor cation channel is blocked by Mg2+ at resting membrane potential. To unblock the channel, the postsynaptic cell must be depolarized.[32]
The NMDA receptor therefore functions as a "molecular coincidence detector". Its ion channel only opens when the following two conditions are met simultaneously: glutamate is bound to the receptor, and the postsynaptic cell is depolarized (which removes the Mg2+ blocking the channel). This property of the NMDA receptor explains many aspects of long term potentiation (LTP) and synaptic plasticity.[33]
NMDA receptors are modulated by a number of endogenous and exogenous compounds and play a key role in a wide range of physiological (e.g. memory) and pathological processes (e.g. excitotoxicity).
Clinical significance
Recently, NMDARs were associated with a rare autoimmune disease, Anti-NMDAR encephalitis, that usually occurs due to cross reactivity of antibodies produced by the immune system against ectopic brain tissues, such as those found in teratoma.[34]
See also
- Long-term potentiation and depression
- Category: NMDA receptor antagonists
- NMDA
- AMPA
- AMPA receptor
- Calcium/calmodulin-dependent protein kinases
- Anti-glutamate receptor antibodies
External links
 Media related to NMDA receptor at Wikimedia Commons
Media related to NMDA receptor at Wikimedia Commons- NMDA receptor pharmacology
- Motor Discoordination Results from Combined Gene Disruption of the NMDA Receptor NR2A and NR2C Subunits, But Not from Single Disruption of the NR2A or NR2C Subunit
- A schematic diagram summarizes three potential models for the switching of NR2A and NR2B subunits at developing synapses. - a figure from Liu et al., 2004[35]
- Drosophila NMDA receptor 1 - The Interactive Fly
References
- ^ Laube, B; Hirai H, Sturgess M, Betz H, and Kuhse J (1997). "Molecular determinants of antagonists discrimination by NMDA receptor subunits: Analysis of the glutamate binding site on the NR2B subunit". Neuron 18 (3): 493–503. doi:10.1016/S0896-6273(00)81249-0. PMID 9115742.
- ^ Clinical Implications of Basic Research: Memory and the NMDA receptors, Fei Li and Joe Z. Tsien, N Engl J Med, 361:302, July 16, 2009
- ^ Dingledine R, Borges K, Bowie D, Traynelis SF (March 1999). "The glutamate receptor ion channels". Pharmacol. Rev. 51 (1): 7–61. PMID 10049997. http://pharmrev.aspetjournals.org/cgi/pmidlookup?view=long&pmid=10049997.
- ^ Liu Y, Zhang J (October 2000). "Recent development in NMDA receptors". Chin. Med. J. 113 (10): 948–56. PMID 11775847. http://www.cmj.org/Periodical/LinkIn.asp?journal=Chinese%20Medical%20Journal&linkintype=pubmed&year=2000&vol=113&issue=10&beginpage=948.
- ^ Cull-Candy S, Brickley S, Farrant M (June 2001). "NMDA receptor subunits: diversity, development and disease". Curr. Opin. Neurobiol. 11 (3): 327–35. doi:10.1016/S0959-4388(00)00215-4. PMID 11399431. http://linkinghub.elsevier.com/retrieve/pii/S0959-4388(00)00215-4.
- ^ Paoletti P, Neyton J (February 2007). "NMDA receptor subunits: function and pharmacology". Curr Opin Pharmacol 7 (1): 39–47. doi:10.1016/j.coph.2006.08.011. PMID 17088105.
- ^ Kleckner, N W; Dingledine, R (1988). "Requirement for glycine in activation of NMDA-receptors expressed in Xenopus oocytes.". Science 241 (4867): 835-837. PMID 2841759. http://www.ncbi.nlm.nih.gov/pubmed/2841759.
- ^ Stephenson FA (November 2006). "Structure and trafficking of NMDA and GABAA receptors". Biochem. Soc. Trans. 34 (Pt 5): 877–81. doi:10.1042/BST0340877. PMID 17052219. http://www.biochemsoctrans.org/bst/034/0877/0340877.pdf.
- ^ Teng, H. J., W.S. Cai, L.L Zhou, J. Zhang, Q. Liu, Y.Q. Wang, W. Dai, M. Zhao, and Z.S. Sun. 2010. Evolutionary Mode and Functional Divergence of Vertebrate NMDA Receptor Subunit 2 Genes. PLoS One 5, doi:ARTN e13342 DOI 10.1371/journal.pone.0013342 (2010).
- ^ Ryan, T. J. & Grant, S. G. N. The origin and evolution of synapses (vol 10, pg 701, 2009). Nat Rev Neurosci 10, doi:Doi 10.1038/Nrn2748 (2009).
- ^ Liu XB, Murray KD, Jones EG (October 2004). "Switching of NMDA receptor 2A and 2B subunits at thalamic and cortical synapses during early postnatal development". J. Neurosci. 24 (40): 8885–95. doi:10.1523/JNEUROSCI.2476-04.2004. PMID 15470155.
- ^ Liu Y, Wong TP, Aarts M, Rooyakkers A, Liu L, Lai TW, Wu DC, Lu J, Tymianski M, Craig AM, Wang YT (March 2007). "NMDA receptor subunits have differential roles in mediating excitotoxic neuronal death both in vitro and in vivo". J. Neurosci. 27 (11): 2846–57. doi:10.1523/JNEUROSCI.0116-07.2007. PMID 17360906.
- ^ Zhou M, Baudry M (March 2006). "Developmental changes in NMDA neurotoxicity reflect developmental changes in subunit composition of NMDA receptors". J. Neurosci. 26 (11): 2956–63. doi:10.1523/JNEUROSCI.4299-05.2006. PMID 16540573.
- ^ Sprengel, R. et al. Importance of the intracellular domain of NR2 subunits for NMDA receptor function in vivo. Cell 92, 279-289 (1998).
- ^ Groc L, Choquet D, Stephenson FA, Verrier D, Manzoni OJ, Chavis P (2007). "NMDA receptor surface trafficking and synaptic subunit composition are developmentally regulated by the extracellular matrix protein Reelin". J. Neurosci. 27 (38): 10165–75. doi:10.1523/JNEUROSCI.1772-07.2007. PMID 17881522.
- ^ Espinosa JS, Luo LJ (March 2008). "Timing neurogenesis and differentiation: insights from quantitative clonal analyses of cerebellar granule cells". J. Neurosci. 28 (10): 2301–12. doi:10.1523/JNEUROSCI.5157-07.2008. PMC 2586640. PMID 18322077. http://www.jneurosci.org/cgi/content/full/28/10/2301.
- ^ Gajendran N, Kapfhammer JP, Lain E, Canepari M, Vogt K, Wisden W,Brenner HR (February 2009). "Neuregulin Signaling Is Dispensable for NMDA- and GABAA-Receptor Expression in the Cerebellum In Vivo". J. Neurosci. 28 (10): 2301–12. doi:10.1523/JNEUROSCI.4303-08.2009. PMID 19244516. http://www.jneurosci.org/cgi/content/full/29/8/2404.
- ^ Chen PE, Geballe MT, Stansfeld PJ, Johnston AR, Yuan H, Jacob AL, Snyder JP, Traynelis SF, Wyllie DJ (May 2005). "Structural features of the glutamate binding site in recombinant NR1/NR2A N-methyl-D-aspartate receptors determined by site-directed mutagenesis and molecular modeling". Mol. Pharmacol. 67 (5): 1470–84. doi:10.1124/mol.104.008185. PMID 15703381.
- ^ Wolosker H (Oct 2006). "D-serine regulation of NMDA receptor activity". Sci. STKE pe41 (356): 1–3. doi:10.1126/stke.3562006pe41. PMID 17033043.
- ^ Anderson C (2003-06-01). "The Bad News Isn't In : A Look at Dissociative-Induced Brain Damage and Cognitive Impairment". Erowid DXM Vaults : Health. http://www.erowid.org/chemicals/dxm/dxm_health2.shtml. Retrieved 2008-12-17.
- ^ "Effects of N-Methyl-D-Aspartate (NMDA)-Receptor Antagonism on Hyperalgesia, Opioid Use, and Pain After Radical Prostatectomy". ClinicalTrials.gov. 2005-09-01. http://www.clinicaltrials.gov/ct/show/NCT00188383?order=4. Retrieved 2008-12-17.
- ^ http://www.clinicalpharmacology-ip.com
- ^ Toxicol. Sci. 2010 116: 249-263; doi:10.1093/toxsci/kfq111
- ^ Huggins DJ, Grant GH (January 2005). "The function of the amino terminal domain in NMDA receptor modulation". J. Mol. Graph. Model. 23 (4): 381–8. doi:10.1016/j.jmgm.2004.11.006. PMID 15670959.
- ^ Eby GA, Eby KL (2006). "Rapid recovery from major depression using magnesium treatment". Medical hypotheses 67 (2): 362–70. doi:10.1016/j.mehy.2006.01.047. PMID 16542786.
- ^ Sucher NJ, Lipton SA (November 1991). "Redox modulatory site of the NMDA receptor-channel complex: regulation by oxidized glutathione". Journal of Neuroscience Research 30 (3): 582–591. doi:10.1002/jnr.490300316. PMID 1666131.
- ^ Aizenman E, Lipton SA, Loring RH (March 1989). "Selective modulation of NMDA responses by reduction and oxidation". Neuron 2 (3): 1257–63. doi:10.1016/0896-6273(89)90310-3. PMID 2696504.
- ^ Yu XM, Askalan R, Keil GJ, Salter MW (January 1997). "NMDA channel regulation by channel-associated protein tyrosine kinase Src". Science 275 (5300): 674–8. doi:10.1126/science.275.5300.674. PMID 9005855.
- ^ Chen Y, Beffert U, Ertunc M, Tang TS, Kavalali ET, Bezprozvanny I, Herz J (September 2005). "Reelin modulates NMDA receptor activity in cortical neurons". J. Neurosci. 25 (36): 8209–16. doi:10.1523/JNEUROSCI.1951-05.2005. PMID 16148228. http://www.jneurosci.org/cgi/pmidlookup?view=long&pmid=16148228.
- ^ Hawasli AH, Benavides DR, Nguyen C, Kansy JW, Hayashi K, Chambon P, Greengard P, Powell CM, Cooper DC, Bibb JA (July 2007). "Cyclin-dependent kinase 5 governs learning and synaptic plasticity via control of NMDAR degradation". Nat. Neurosci. 10 (7): 880–6. doi:10.1038/nn1914. PMID 17529984.
- ^ Zhang S, Edelmann L, Liu J, Crandall JE, Morabito MA (January 2008). "Cdk5 regulates the phosphorylation of tyrosine 1472 NR2B and the surface expression of NMDA receptors". J. Neurosci. 28 (2): 415–24. doi:10.1523/JNEUROSCI.1900-07.2008. PMID 18184784. http://www.jneurosci.org/cgi/pmidlookup?view=long&pmid=18184784.
- ^ Purves, Dale; George J. Augustine, David Fitzpatrick, William C. Hall, Anthony-Samuel LaMantia, James O. McNamara, Leonard E. White (2008). Neuroscience, 4th Ed.. Sinauer Associates. pp. 129–131. ISBN 978-0-87893-697-7. http://www.sinauer.com/neuroscience4e.
- ^ Purves, Dale; George J. Augustine, David Fitzpatrick, William C. Hall, Anthony-Samuel LaMantia, James O. McNamara, Leonard E. White (2008). Neuroscience, 4th Ed.. Sinauer Associates. pp. 191–195. ISBN 978-0-87893-697-7. http://www.sinauer.com/neuroscience4e.
- ^ Iizuka T, Sakai F (September 2008). "[Anti-nMDA receptor encephalitis--clinical manifestations and pathophysiology]" (in Japanese). Brain Nerve 60 (9): 1047–60. PMID 18807939.
- ^ Liu XB, Murray KD, Jones EG.(2004) Switching of NMDA receptor 2A and 2B subunits at thalamic and cortical synapses during early postnatal development. The Journal of Neuroscience, 24(40):8885-95.PMID 15470155 free fulltext
Ion channel, cell surface receptor: ligand-gated ion channels Cys-loop receptors monomers: α1 · α2 · α3 · α4 · α5 · α6 · α7 · α9 · α10 · β1 · β2 · β3 · β4 · δ · ε
pentamers: (α3)2(β4)3 · (α4)2(β2)3 · (α7)5 · (α1)2(β4)3 - Ganglion type · (α1)2β1δε - Muscle typeZincZinc-activatedIonotropic glutamates Ligand-gated onlyVoltage- and ligand-gated‘Orphan’ATP-gated channels B trdu: iter (nrpl/grfl/cytl/horl), csrc (lgic, enzr, gprc, igsr, intg, nrpr/grfr/cytr), itra (adap, gbpr, mapk), calc, lipd; path (hedp, wntp, tgfp+mapp, notp, jakp, fsap, hipp, tlrp) Neuroethology Concepts Feedforward · Coincidence detector · Umwelt · Instinct · Feature detection · Central pattern generator (CPG) · NMDA receptor · Lateral inhibition · Fixed action pattern · Krogh's Principle · Hebbian theory · Anti-Hebbian learning · Sound localization · Ultrasound avoidance in insects ·
People Theodore Holmes Bullock · Walter Heiligenberg · Niko Tinbergen · Konrad Lorenz · Donald Griffin · Donald Kennedy · Karl von Frisch · Erich von Holst · Jörg-Peter Ewert · Franz Huber · Bernhard Hassenstein · Werner E. Reichardt · Eric Knudsen · Eric Kandel · Nobuo Suga · Masakazu Konishi · Fernando Nottebohm
Methods Whole Cell Patch Clamp · Slice Preparation
Systems Animal echolocation · Waggle dance · Jamming avoidance response · Vision in toads · Frog hearing and communication · Infrared sensing in snakes · Caridoid escape reaction · Vocal learning
Anxiety disorder: Obsessive–compulsive disorder (F42, 300.3) History Yale–Brown Obsessive Compulsive ScaleBiology NeuroanatomyReceptorsSymptoms Obsessions (associative, diagnostic, injurious, scrupulous, pathogenic, sexual) · Compulsions (impulses, rituals, tics) · Thought suppression (avoidance) · Hoarding (animals, books, possessions)Treatment Mu opioidergicsNMDA glutamatergicsNK-1 tachykininergicsOtherBehavioralOrganizations Notable people Edna B. Foa · Stanley Rachman · Adam S. Radomsky · Jeffrey M. Schwartz · Susan Swedo · Jeff Bell · Emily ColasPopular culture LiteratureFictionalNonfictionEverything in Its Place · Just CheckingMediaRelated Obsessive–compulsive personality disorder · Obsessional jealousy · Purely Obsessional OCD · Social anxiety disorder · Tourette syndromeCategories:- Ionotropic receptors
- Ion channels
- Molecular neuroscience
- Cell signaling


Wikimedia Foundation. 2010.