- Lateral geniculate nucleus
-
Brain: Lateral geniculate nucleus 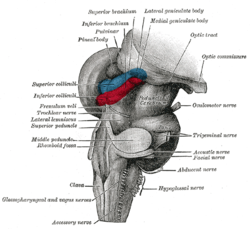
Hind- and mid-brains; postero-lateral view. (Lateral geniculate body visible near top.) Latin Corpus geniculatum laterale Part of Thalamus System Visual Artery Anterior choroidal and Posterior cerebral Vein Terminal vein NeuroNames hier-335 NeuroLex ID birnlex_1662 The lateral geniculate nucleus (LGN) is the primary relay center for visual information received from the retina of the eye. The LGN is found inside the thalamus of the brain.
The LGN receives information directly from the ascending retinal ganglion cells via the optic tract and from the reticular activating system. Neurons of the LGN send their axons through the optic radiation, a pathway directly to the primary visual cortex. In addition, the LGN receives many strong feedback connections from the primary visual cortex. In mammals and humans the two strongest pathways linking the eye to the brain are those projecting to the LGNd (dorsal part of the LGN in the thalamus), and to the Superior Colliculus (SC)[1]
Contents
Structure
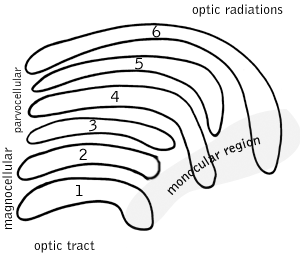
Both the left and right hemisphere of the brain have a lateral geniculate nucleus, named so for its resemblance to a bent knee (genu is Latin for "knee"). In many primates, including humans and macaques, it has layers of cell bodies with layers of neuropil in between, in an arrangement something like a club sandwich or layer cake, with cell bodies of LGN neurons as the "cake" and neuropil as the "icing". In humans and macaques the LGN is normally described as having six distinctive layers. The inner two layers, 1 and 2, are called the magnocellular layers, while the outer four layers, 3, 4, 5, and 6, are called parvocellular layers. An additional set of neurons, known as the koniocellular sublayers, are found ventral to each of the magnocellular and parvocellular layers.[2] It must be noted, this layering is variable between primate species, and extra leafleting is variable within species.
M, P, K cells
Type Size* Source / Type of Information Location Response Number M: Magnocellular cells Large Rods; necessary for the perception of movement, depth, and small differences in brightness Layers 1 and 2 rapid and transient 95% (120 Mio) P: Parvocellular cells (or "parvicellular") Small Cones; long- and medium-wavelength ("red" and "green" cones); necessary for the perception of color and form (fine details). Layers 3, 4, 5 and 6 slow and sustained 5% (6 Mio) K: Koniocellular cells (or "interlaminar") Very small cell bodies Short-wavelength "blue" cones. Between each of the M and P layers ? 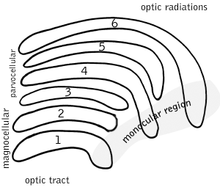 Schematic diagram of the primate LGN. Layers 1 and 2 are more ventrally located, and are next to the incoming optic tract fibers.
Schematic diagram of the primate LGN. Layers 1 and 2 are more ventrally located, and are next to the incoming optic tract fibers.
- Size relates to cell body, dendritic tree and receptive field
The magnocellular, parvocellular, and koniocellular layers of the LGN correspond with the similarly named types of ganglion cells.
Koniocellular cells are functionally and neurochemically distinct from M and P cells and provide a third channel to the visual cortex. They project their axons between the layers of the lateral geniculate nucleus where M and P cells project. Their role in visual perception is presently unclear; however, the koniocellular system has been linked with the integration of somatosensory system-proprioceptive information with visual perception, and it may also be involved in color perception.[citation needed]
The parvo- and magnocellular fibers were previously thought to dominate the Ungerleider-Mishkin ventral stream and dorsal stream, respectively. However, new evidence has accumulated showing that the two streams appear to feed on a more even mixture of different types of nerve fibers.[3]
The other major retino-cortical visual pathway is the retinotectal pathway, routing primarily through the superior colliculus and thalamic pulvinar nucleus onto posterior parietal cortex and visual area MT.
Ipsilateral and contralateral layers
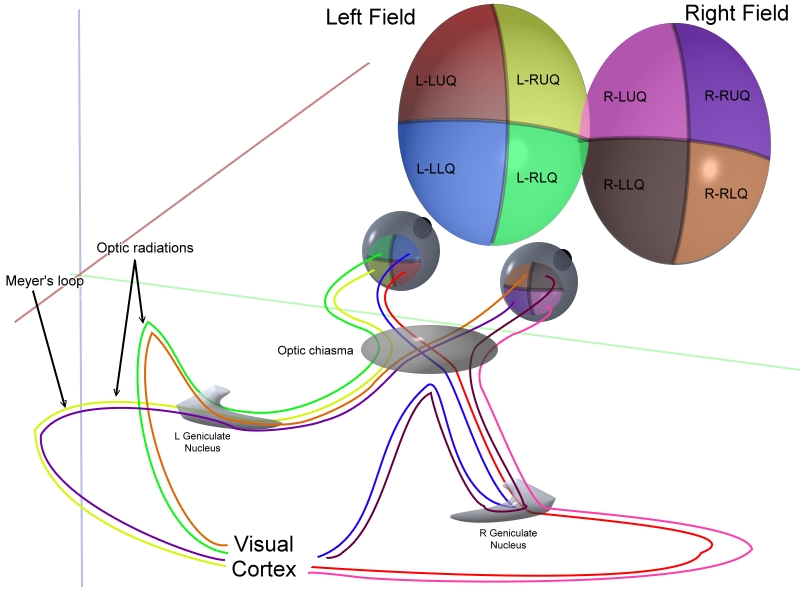
Both the LGN in the right hemisphere and the LGN in the left hemisphere receive input from each eye. However, each LGN only receives information from one half of the visual field. This occurs due to axons of the ganglion cells from the inner halves of the retina (the nasal sides) decussating (crossing to the other side of the brain) through the optic chiasm (khiasma means "cross"). The axons of the ganglion cells from the outer half of the retina (the temporal sides) remain on the same side of the brain. Therefore, the right hemisphere receives visual information from the left visual field, and the left hemisphere receives visual information from the right visual field. Within one LGN, the visual information is divided among the various layers as follows:[4]
- the eye on the same side (the ipsilateral eye) sends information to layers 2, 3 and 5
- the eye on the opposite side (the contralateral eye) sends information to layers 1, 4 and 6.
A simple mnemonic for remembering this is "See I? I see, I see," with "see" representing the C in "contralateral," and "I" representing the I in "ipsilateral."
Another way of remembering this is 2+3=5, which is correct, so ipsilateral side, and 1+4 doesn't equal 6, so contralateral.
This description applies to the LGN of many primates, but not all. The sequence of layers receiving information from the ipsilateral and contralateral (opposite side of the head) eyes is different in the tarsier.[5] Some neuroscientists suggested that "this apparent difference distinguishes tarsiers from all other primates, reinforcing the view that they arose in an early, independent line of primate evolution".[6]
In visual perception, the right eye gets information from the right side of the world (the right visual field), as well as the left side of the world (the left visual field). You can confirm this by covering your left eye: the right eye still sees to your left and right, although on the left side your field of view is partially blocked by your nose.
In the LGN, the corresponding information from the right and left eyes is "stacked" so that a toothpick driven through the club sandwich of layers 1 through 6 would hit the same point in visual space six different times.
LGN inputs
The LGN receives input from many sources, including the cortex and then sends its output to the cortex.
At least in some species, the LGN also receives some inputs from the optic tectum (also known as the superior colliculus).[7]
LGN output
Information leaving the LGN travels out on the optic radiations, which form part of the retrolenticular limb of the internal capsule.
The axons that leave the LGN go to V1 visual cortex. Both the magnocellular layers 1-2 and the parvocellular layers 3-6 send their axons to layer 4 in V1. Within layer 4 of V1, layer 4cβ receives parvocellular input, and layer 4cα receives magnocellular input. However, the koniocellular layers (in between layers 1-6) send their axons to layers 4a in V1. Axons from layer 6 of visual cortex send information back to the LGN.
Studies involving blindsight have suggested that projections from the LGN not only travel to the primary visual cortex but also to higher cortical areas V2 and V3. Patients with blindsight are phenomenally blind in certain areas of the visual field corresponding to a contralateral lesion in primary visual cortex; however, these patients are able to perform certain motor tasks accurately in their blind field, such as grasping. This suggests that neurons travel from the LGN to both the visual cortex and higher cortex regions.[8]
Function in visual perception
The function of the LGN is unknown. It has been shown that while the retina accomplishes spatial decorrelation through center surround inhibition, the LGN accomplishes temporal decorrelation.[9] This spatial-temporal decorrelation makes for much more efficient coding. However, there is almost certainly much more going on.
Like other areas of the thalamus, particularly other relay nuclei, the LGN likely helps the visual system focus its attention on the most important information. That is, if you hear a sound slightly to your left, the auditory system likely "tells" the visual system, through the LGN, to direct visual attention to that part of space.
The LGN is also a station that refines certain receptive fields.
Recent experiments using fMRI in humans have found that both spatial attention and saccadic eye movements can modulate activity in the LGN.
Additional images
-
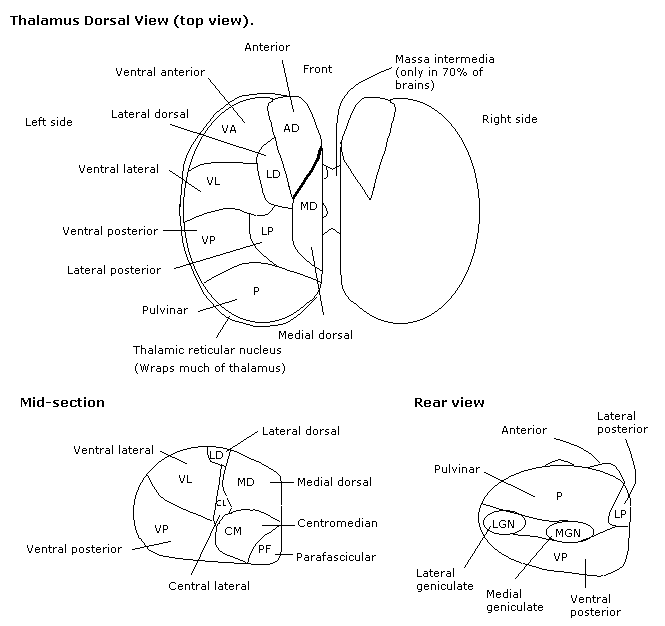
Thalamus
-
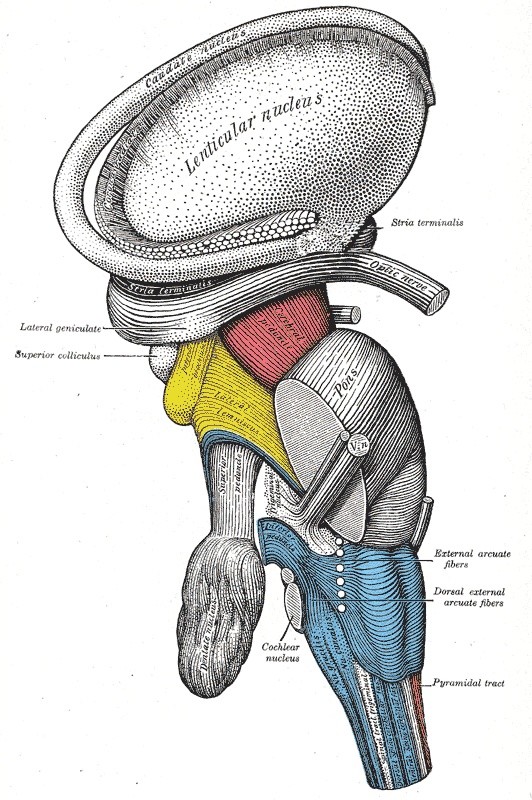
Dissection of brain-stem. Lateral view.
-
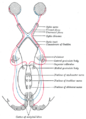
Scheme showing central connections of the optic nerves and optic tracts.
-
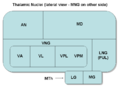
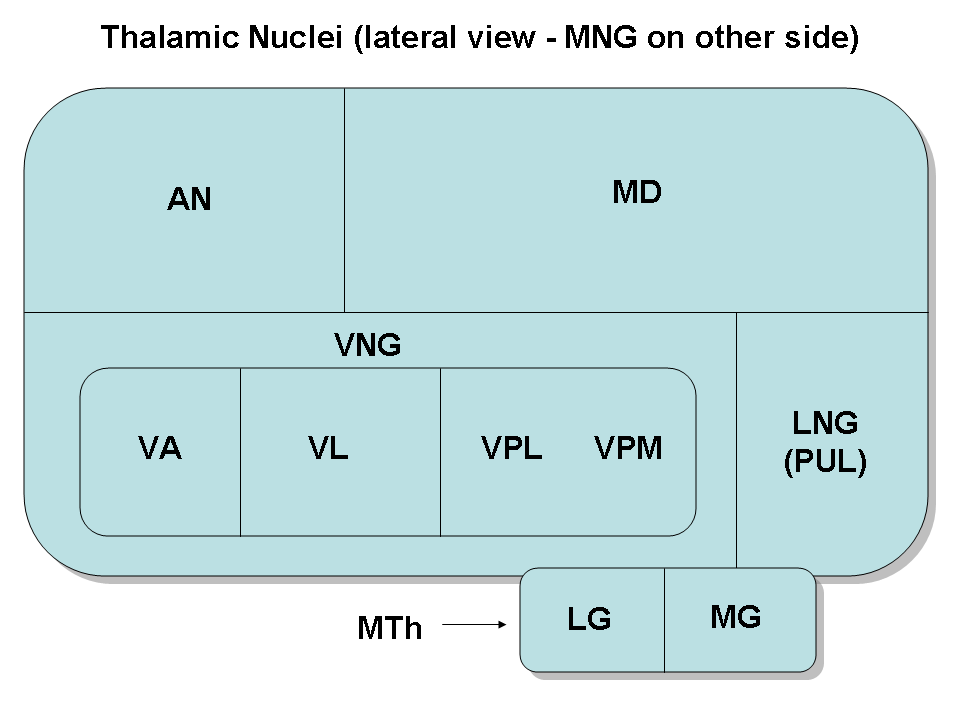
Thalamic nuclei
-
3D schematic representation of optic tracts
References
- ^ Goodale, M. & Milner, D. (2004)Sight unseen.Oxford University Press, Inc.: New York.
- ^ Carlson, N. R. (2007)Physiology of Behavior: ninth edition.Pearson Education, Inc.: Boston.
- ^ Goodale & Milner, 1993, 1995.
- ^ Nicholls J., et al. From Neuron to Brain: Fourth Edition. Sinauer Associates, Inc. 2001.
- ^ Rosa MG, Pettigrew JD, Cooper HM (1996) Unusual pattern of retinogeniculate projections in the controversial primate Tarsius. Brain Behav Evol 48(3):121-129.
- ^ Collins CE, Hendrickson A, Kaas JH (2005) Overview of the visual system of Tarsius. Anat Rec A Discov Mol Cell Evol Biol 287(1):1013-1025.
- ^ In Chapter 7, section "The Parcellation Hypothesis" of "Principles of Brain Evolution", Georg F. Striedter (Sinauer Associates, Sunderland, MA, USA, 2005) states, "...we now know that the LGN receives at least some inputs from the optic tectum (or superior colliculus) in many amniotes". He cites "Wild, J.M. 1989. Pretectal and tectal projections to the homolog of the dorsal lateral geniculate nucleus in the pigeon - an anterograde and retrograde tracing study with cholera-toxin conjugated to horseradish-peroxidase. Brain Res 489: 130-137" and also "Kaas, J.H., and Huerta, M.F. 1988. The subcortical visual system of primates. In: Steklis H. D., Erwin J., editors. Comparative primate biology, vol 4: neurosciences. New York: Alan Liss, pp. 327-391.
- ^ Schmid, Michael C.; Mrowka, Sylwia W.; Turchi, Janita et al. (2010). "Blindsight depends on the lateral geniculate nucleus". Nature 466 (7304): 373–377. doi:10.1038/nature09179.
- ^ Dawei W. Dong and Joseph J. Atick, Network - Temporal Decorrelation: A Theory of Lagged and Nonlagged Responses in the Lateral Geniculate Nucleus, 1995, pp. 159-178.
External links
- Malpeli J. Malpeli Lab Home Page. Retrieved September 1, 2004.
- BrainMaps at UCDavis lateral%20geniculate%20nucleus
- Atlas of anatomy at UMich eye_38 - "The Visual Pathway from Below"
- BrainMaps at UCDavis lgn
Sensory system: Visual system and eye movement pathways Visual perception 1° (Bipolar cell of Retina) → 2° (Ganglionic cell) → 3° (Optic nerve → Optic chiasm → Optic tract → LGN of Thalamus) → 4° (Optic radiation → Cuneus and Lingual gyrus of Visual cortex → Blobs → Globs)Muscles of orbit TrackingHorizontal gazeVertical gazePupillary reflex Pupillary dilation1° (Posterior hypothalamus → Ciliospinal center) → 2° (Superior cervical ganglion) → 3° (Sympathetic root of ciliary ganglion → Nasociliary nerve → Long ciliary nerves → Iris dilator muscle)1° (Retina → Optic nerve → Optic chiasm → Optic tract → Visual cortex → Brodmann area 19 → Pretectal area) → 2° (Edinger-Westphal nucleus) → 3° (Short ciliary nerves → Ciliary ganglion → Ciliary muscle)Circadian rhythm M: EYE
anat(g/a/p)/phys/devp/prot
noco/cong/tumr, epon
proc, drug(S1A/1E/1F/1L)
Categories:


Wikimedia Foundation. 2010.